松江鲈白介素15(TfIL-15)的结构特征与重组表达*
2019-08-05刘莹莹于珊珊柴迎梅林啸鹏
刘莹莹 于珊珊 柴迎梅 林啸鹏 祝 茜
松江鲈白介素15(TfIL-15)的结构特征与重组表达*
刘莹莹 于珊珊 柴迎梅 林啸鹏 祝 茜①
(山东大学(威海)海洋学院 威海 264209)
为研究白介素15(Interleukin-15, IL-15)在松江鲈(Heckel)先天免疫中的功能,本研究利用RACE技术克隆得到松江鲈IL-15基因(命名为TfIL-15)的全长cDNA序列,其长度为1140 bp,包括5¢-非编码区(5¢-UTR) 165 bp、开放阅读框(ORF) 522 bp和3¢UTR 453 bp。在5¢UTR区域,存在4个读码框外的AUG翻译起始位点。基因ORF编码173个氨基酸(aa),其中,前59 aa为信号肽序列。成熟肽全长为114 aa,预测分子量为12.975 kDa,理论等电点为5.15。同源比对发现,鱼类IL-15变异程度较高,TfIL-15与其他鱼类IL-15同源性在23%~61%之间。多序列比对和三维结构构建结果显示,TfIL-15具有典型4个α螺旋二级结构,形成二硫键的4个半胱氨酸高度保守。qRT-PCR分析表明,TfIL-15广泛表达于松江鲈各组织中。腹腔注射脂多糖(Lipopolysaccharides, LPS)后,TfIL-15 mRNA在血液、皮肤、肝脏和脾脏中均上调表达。在皮肤和血液中,刺激后2 h表达量迅速上调至最高峰,分别为对照组的74倍和41倍。脾脏和肝脏在刺激后12 h分别达到对照组的3倍和18倍。肝脏中,刺激后96 h,表达量再次上调至对照组的86倍。上述结果表明,TfIL-15可能参与了松江鲈抵抗外界刺激的先天免疫过程。另外,通过构建TfIL-15成熟肽的原核表达载体,成功获得重组蛋白,为进一步研究TfIL-15蛋白的功能奠定了基础。
松江鲈;白介素15(IL-15);克隆;基因表达;重组蛋白表达
IL-15是白介素(Interleukin, IL)家族的重要成员之一,它与IL-2、GM-CSF(粒细胞-巨噬细胞集落刺激因子)、G-CSF(粒细胞集落刺激因子)等,同属于4α螺旋细胞因子家族(Lodolce, 2002)。转录水平的IL-15存在于多种类型细胞和组织器官中,包括表皮细胞、上皮细胞、神经细胞、单核细胞、巨噬细胞、树突状细胞及心脏、肺、脾和骨骼肌等(Carson, 1995; Lee, 1996)。但蛋白水平的IL-15在血清或培养细胞的上清液中却很难被检测到。这种现象与IL-15基因的蛋白表达受严谨调控相关,这些调控方式包括5¢-UTR多个翻译起始位的存在、特殊的信号肽及羧基末端的某些结构等(Bamford, 1996、1998; Kurys, 2000)。IL-15是一种多效细胞因子,具有增强淋巴细胞活性、促进粒细胞吞噬(Ratthé, 2004)和NK细胞发育并增强其细胞毒性的作用(Fehniger, 2001; Lodolce, 2002)。此外,IL-15还具有刺激T细胞/B细胞增殖、诱导免疫球蛋白分泌、刺激特异性抗原产生、诱导信号转导和转录激活蛋白表达等多种功能(王东勇等, 1996)。
尽管功能多样,但由于白介素等细胞因子在趋异进化过程中的变异程度相当高,导致鱼类相关基因的开发难度加大,免疫功能研究相对滞后(贝锦新, 2006)。2004年之前,硬骨鱼IL-15的相关研究工作几乎未见报道。随着红鳍东方鲀()、黑青斑河豚()和斑马鱼()等物种基因组计划的完成,一系列细胞因子包括IL-15才开始被陆续发现和鉴定(Bei, 2006; Fang, 2006; Gunimaladevi, 2007)。除了以上3种鱼,目前,lL-15也仅在虹鳟()、三刺鱼()、金头鲷()、条石鲷()、露斯塔野鲮(, Hamilton)等少数鱼类中克隆得到(方玮, 2009; Das, 2015; Pérez-Cordón, 2014; Wang, 2007)。研究发现,鱼类lL-15可以由多种组织和细胞产生,组织分布广泛;受到不同诱导物免疫刺激后,lL-15表达上调,但上调时间和不同组织的表现有所不同(贝锦新, 2006;方玮, 2009)。此外,lL-15还参与了鱼体对抗寄生虫的免疫反应(Pérez-Cordón, 2014; Das, 2015)。虹鳟体外重组表达lL-15蛋白能诱导虹鳟脾脏细胞产生IFN-γ (Wang, 2007)。红鳍东方鲀重组表达的lL-15,能活化鱼的外周血白细胞、小鼠T淋巴细胞(CTLL-2)、鱼的头肾细胞和胸腺细胞,说明重组蛋白具有活化增殖免疫细胞的生物学功能(贝锦新, 2006;孙赛红等, 2015)。这些研究为今后进一步深入开展重组lL-15在鱼类健康养殖上的应用研究提供了基础。
松江鲈(Heckel),隶属于鲉形目(Scorpaeniformes)、杜父鱼科(Cottidae)、松江鲈属(),是一种近海溯河洄游的肉食性鱼类(于诗群等, 2008; 陈学昭等, 2016)。近年来,由于种群数量剧减,被列为国家Ⅱ级保护动物(陈学昭等, 2016)。目前,关于IL-15基因在松江鲈体内的研究尚未开展,本研究克隆了松江鲈IL-15(命名为TfIL-15)的基因,分析了序列结构特征,应用实时荧光定量PCR(Quantitative real-time PCR, qRT-PCR)技术研究了其在鱼体内的分布和免疫刺激后基因应答情况,并利用原核表达系统体外表达了IL-15成熟肽的重组蛋白。研究结果可进一步丰富鱼类细胞因子的研究内容,有利于阐明鱼类细胞因子与鱼类免疫系统的关系。重组蛋白的表达为进一步开展对TfIL-15的功能研究和抗体制备奠定了基础。
1 材料与方法
1.1 材料与组织样品制备
9~10月龄的松江鲈(体重约15~23 g)取自山东文登埠口松江鲈自然保护区,实验前置于充气海水中(12℃~14℃)饲养1周。选取正常健康个体,麻醉后心脏取血,然后解剖取其心脏、肝脏、鳃、肠、皮肤、肾脏、脾脏、脑、卵等组织,立即放入Trizol中研磨,用于提取正常组织RNA。
LPS (Lipopolysaccharides, 脂多糖)刺激实验,将松江鲈平均分成2组(每组50条),一组腹腔注50 μl (0.04 mg/kg)的LPS,对照组注射等体积无菌生理盐水。刺激后0、2、6、12、24、48、72、96 h麻醉取样(每次6条),分别取其血、皮肤、肝脏和脾脏用于组织总RNA提取。
1.2 总RNA的提取和cDNA合成
依据Trizol (Invitrogen公司)操作说明提取各样品的组织总mRNA。cDNA第一链的合成参照Clontech公司SMART(Switching mechanism at 5¢end of RNA template)的指导说明进行(表1)(史晓丽等, 2018)。
1.3 TfIL-15 cDNA的全长克隆
根据LPS刺激后cDNA文库的构建获得TfIL-15基因的EST序列,利用Primer 5.0设计特异性后引物15R与5¢primer配对进行5¢RACE克隆;设计特异性前引物15F与3¢anchor R配对进行3¢RACE克隆。所用模板为松江鲈刺激后肝脏cDNA,反应条件:94℃预变性5 min;94℃变性30 s,55℃复性30 s,72℃延伸40 s,30个循环;72℃延伸7 min。然后将PCR纯化产物与pMD-18T载体连接,克隆测序,得到5¢端和3¢端序列片段。
1.4 TfIL-15基因的序列分析
利用软件BioEdit对RACE获得的两端cDAN序列进行拼接,以获得TfIL-15的cDNA全长序列;通过在线软件ExPASy(http://www.au.expasy.org/)对TfIL- 15的cDNA序列进行蛋白翻译、等电点及分子量预测;TfIL-15编码蛋白的信号肽、糖基化位点分别应用SignalIP 4.1和NetNGlyc1.0 Server进行分析;蛋白的三级结构预测在SWISS-MODEL (http://swissmodel. expasy.org/)进行,然后用PyMol软件进行三维结构的建构与标注;在NCBI下载同源序列,利用ClustalW、BioEdit和DNAman进行同源序列比对分析,利用MEGA5.0软件以邻位法(Neighbor-joining, NJ)构建系统发育树。
1.5 组织表达分析
TfIL-15的组成型表达及刺激后表达模式的变化通过qRT-PCR分析来进行。根据TfIL-15 cDNA序列设计qRT-PCR引物(表1),以正常各组织及刺激组和对照组的不同时间点cDNA为模板,进行反应。反应条件:94℃ 3 min;94℃ 15 s,60℃ 60 s,40个循环(陈学昭等, 2016),结束后进行熔解曲线分析。以β-actin作为内参基因,每个样品cDNA设3个平行实验。反应和分析使用7300实时荧光定量PCR仪(Applied Biosystems, 美国)进行。基因的相对表达量根据2-ΔΔCt法计算,结果用平均值±标准差(Mean±SD)表示(马慧鑫等, 2018)。差异显著性使用T检验的 方法分析,当差异显著性水平<0.05时认为差异 显著。
1.6 TfIL-15重组蛋白表达
TfIL-15基因成熟肽的两端引物分别引入Ⅰ和Ⅰ酶切位点,插入到pET-30a(+)表达载体的多克隆位点中,构建pET-30a(+)/TfIL-15表达载体转化入大肠杆菌克隆菌株DH5α中。经阳性克隆筛选后,提取构建好的质粒载体转化表达菌株BL21(DE3)并进行阳性筛选。
将筛选出的菌落分别置于LB液体培养基中过夜培养。次日,将过夜菌转培养(1 : 100的比例)至新的液体LB培养基中约3 h。待其OD600 nm为0.6~0.8时,加入终浓度为0.5 mmol/L的IPTG进行诱导。诱导结束后,收集菌体,通过SDS-PAGE检测目的蛋白的表达情况。进一步的超声破碎检测发现,蛋白以包涵体形式表达。
表1 PCR引物名称及序列

Tab.1 PCR primers and the sequences
在确定重组蛋白表达的情况下,进行蛋白的大规模培养表达,表达结束用PBS重悬菌体超声破碎1 h。将超声破碎后沉淀进行包涵体洗涤,然后用GenScript的His-tag亲和层析柱进行纯化,具体操作步骤参见Yu等(2013)的方法。重组蛋白的复性采用尿素梯度透析法,每个梯度4℃透析16 h (王彤等, 2013)。透析结束后,通过SDS-PAGE检测重组蛋白的纯化结果。
2 结果
2.1 TfIL-15基因克隆及序列分析
松江鲈IL-15全长cDNA序列(图1,GenBank登录号为MH299805)为1140 bp,其中,5¢-非编码区(5¢-UTR) 165 bp,开放阅读框(ORF)522 bp,3¢-UTR 453 bp。在5¢-UTR区域,存在4个读码框外的AUG翻译起始位点。基因ORF编码173个氨基酸(aa),其中,前59 aa为信号肽序列。成熟肽全长为114 aa,预测分子量为12.975 kDa,理论等电点为5.15。成熟肽含有2个可能的N糖基化位点:NASM,132~134;NITV,153~155。
将TfIL-15与其他物种同源序列进行比对,发现TfIL-15与条石鲷(BAN84544.1)的同源性最高为61%,其次为三刺鱼(NP_001254613.1) 59%,与斑马鱼同源性为23%,与鱼类序列同源性在23%~61%之间。与鸡(, AAF61446.1)同源性为24%,与人类IL-15(, AAA21551.1)同源性为25%,与哺乳类同源性在20%~25%之间(表2)。氨基酸序列多序列比对发现,鱼类IL-15与鸟类和哺乳类IL-15的成熟肽区域的同源性较信号肽高,鱼类lL-15同源序列含有与鸟类和哺乳类相同的4个保守的半胱氨酸残基(图2)。

图1 TfIL-15的cDNA序列及其编码序列
下划线标出的分别为起始密码子和终止密码子。阴影部分为N-糖基化位点。箭头表示信号肽切割位点。方框内为5¢-UTR区读码框外的ATG翻译起始位点
The start codon and stop codon are underlined, the N-glycosylation sites are shadowed by gray, the arrow indicates the signal peptide cleavage site, and the four out-of-frame ATG initiation codons in the 5¢UTR are indicated in the boxes
用SWISS-MODEL软件进行三维结构预测,发现TfIL-15具有典型4个α螺旋二级结构(图3A)。将TfIL-15与人类的IL-15叠合时(图3B),发现蛋白结构较吻合,并且二硫键形成位点相互重合。此外,松江鲈另外2个半胱氨酸形成了第3个二硫键(图3A),TfIL-15成熟肽起始氨基酸丙氨酸(A)与人类成熟肽起始氨基酸天冬酰胺(N)重合。
系统进化分析表明,所有鱼类的IL-15聚为一大支,鸟类和哺乳动物的IL-15聚为另外一大支(图4)。
2.2 TfIL-15组织表达分析
qRT-PCR结果表明,TfIL-15 mRNA广泛表达于松江鲈各组织中。其中,在心脏中的表达量最高,其次为鳃(图5)。
2.3 LPS刺激后各组织中TfIL-15基因的表达变化
腹腔注射LPS后,TfIL-15 mRNA在血液、皮肤、肝脏和脾脏中均呈上调表达(图6)。在皮肤中,刺激后2 h表达量迅速上调至最高峰,为对照组的74倍,之后一直呈现较高的表达水平直至刺激后96 h;在血液中,刺激后2 h表达量上调至对照组的41倍,6 h表达量下降至对照组的0.23倍,之后表达量一直显著低于对照组直至实验结束;在脾脏中,刺激后2 h表达量开始上调至对照组的1.3倍,6 h略有下调,12 h达到对照组的3倍左右,之后恢复至对照组水平。在肝脏中,刺激后表达量逐渐上调,2 h、6 h表达量分别上调至对照组的2.2倍和2.4倍,12 h达到对照组的18倍,后恢复对照组水平;但在刺激后96 h,表达量再次上调至在对照组的86倍。
表2 脊椎动物IL-15同源性比对分析
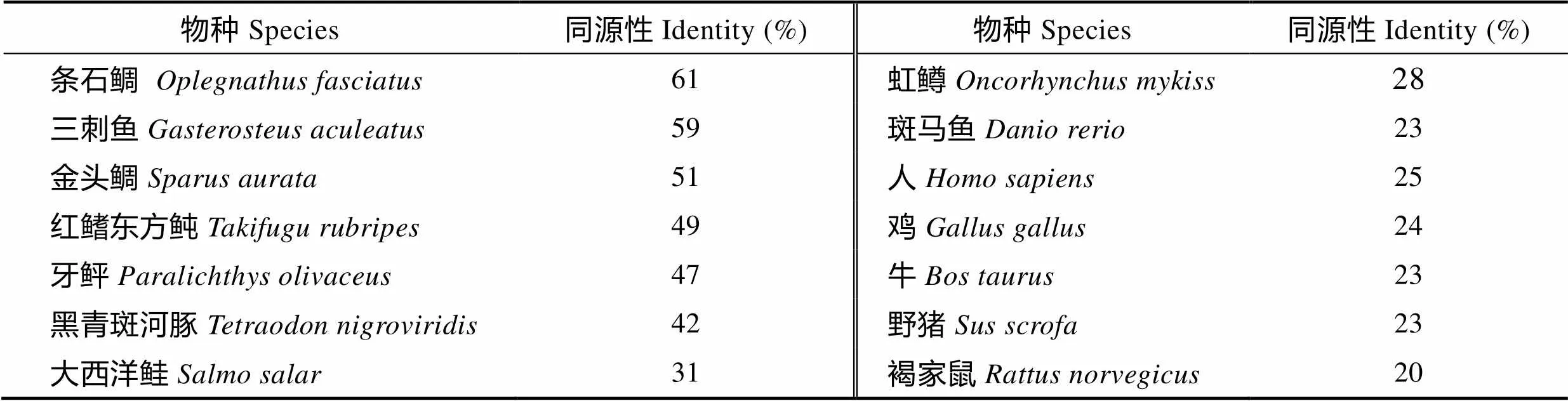
Tab.2 Amino acid identity of TfIL-15 with other vertebrate IL-15s

图2 不同物种IL-15氨基酸序列的多序列比对
保守的半胱氨酸以方框表示,箭头表示形成的二硫键结构,信号肽切割位点以]表示。序列的GenBank登录号分别为:人(AAA21551.1), 褐家鼠(AAB94536.1), 牛(AAA85130.1), 野猪(ABI81495.1), 鸡(AAF61446.1), 黑青斑河豚(AAR25702.1), 红鳍东方鲀(CAF28987.2), 金头鲷(AGS55349.1), 大西洋鲑(AFB81536.1), 虹鳟鱼(CAD88594.1)
The letters in box are the four conserved cysteine residues, the arrows indicate the formed two potential disulphide bonds, and semi square bracket indicates the potential signal peptide cleavage site
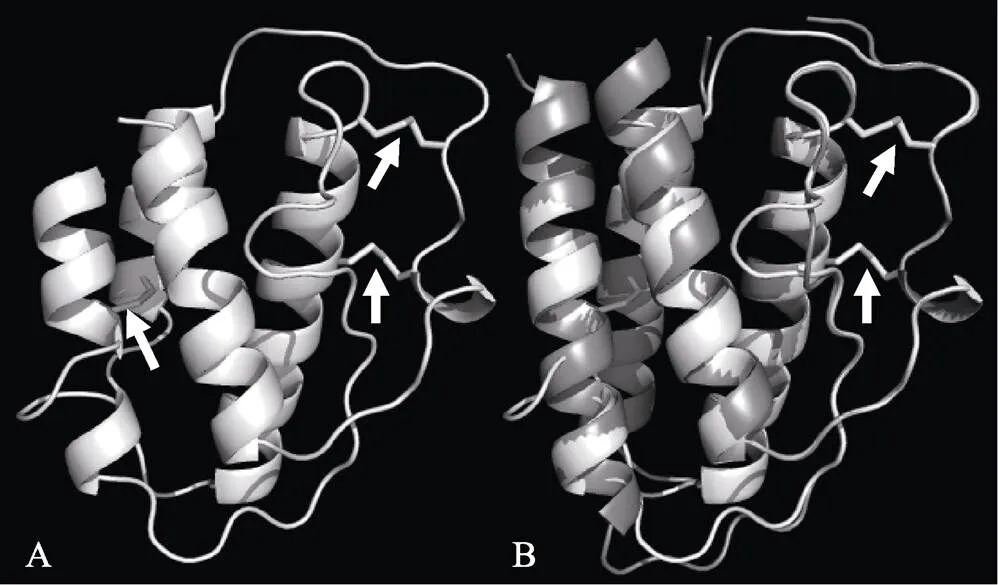
图3 理论预测的TfIL-15蛋白(A)及与人类IL-15(灰色)结构叠合图(白色) (B)
二硫键位置以白色箭头表示
The disulfides are indicated with arrows

图4 基于不同物种IL-15的氨基酸序列构建的系统发育树
三刺鱼的GenBank登录号为NP_001254613.1,其余物种的GenBank登录号同图2,星号表示本研究物种
GenBank accession number ofis NP_001254613.1, and the others accession numbers are the same as in Fig.2. The position of TfIL-15 is marked by an asterisk

图5 TfIL-15基因在正常组织中的表达模式
2.4 TfIL-15的原核表达及纯化
将构建的TfIL-15原核表达菌株经IPTG诱导后进行SDS-PAGE,在19 kDa处出现一条高表达蛋白的特异性条带,与预期大小相符(图7)。将以包涵体形式表达的重组蛋白经亲和层析柱纯化、尿素梯度透析复性,可用于后续的活性检测。
3 讨论
本研究成功克隆到松江鲈IL-15基因cDNA全长序列,跟其他鱼类、哺乳类和鸟类的IL-15基因一样,TfIL-15的5¢-UTR含有多个(4个)读码框外的AUG翻译起始位点(红鳍东方鲀2个,斑马鱼4个,黑青斑河豚2个,虹鳟7个)。TfIL-15含有一段很长的信号肽序列,这也与其他鱼类(松江鲈59 aa,红鳍东方鲀 53 aa,斑马鱼68 aa,虹鳟65 aa,黑青斑河豚53 aa) 以及哺乳类和鸟类相似(人类含有48 aa信号肽,鸟类66 aa)。脊椎动物IL-15这种现象的存在可能与IL-15的转录后表达受到严谨的调控有关(Bamford, 1996、1998; Kurys, 2000)。TfIL-15与条石鲷IL-15的同源性最高,尽管如此,也只有61%。与鱼类同源性在23%~61%之间,可以看出不同物种鱼类IL-15变异程度比较大,这可能是为了适应不同的生长环境而进化所致(TfIL-15与同为海水或者咸淡水鱼类同源性高于淡水鱼类),同时这可能也是目前鱼类IL-15研究较少的原因之一。
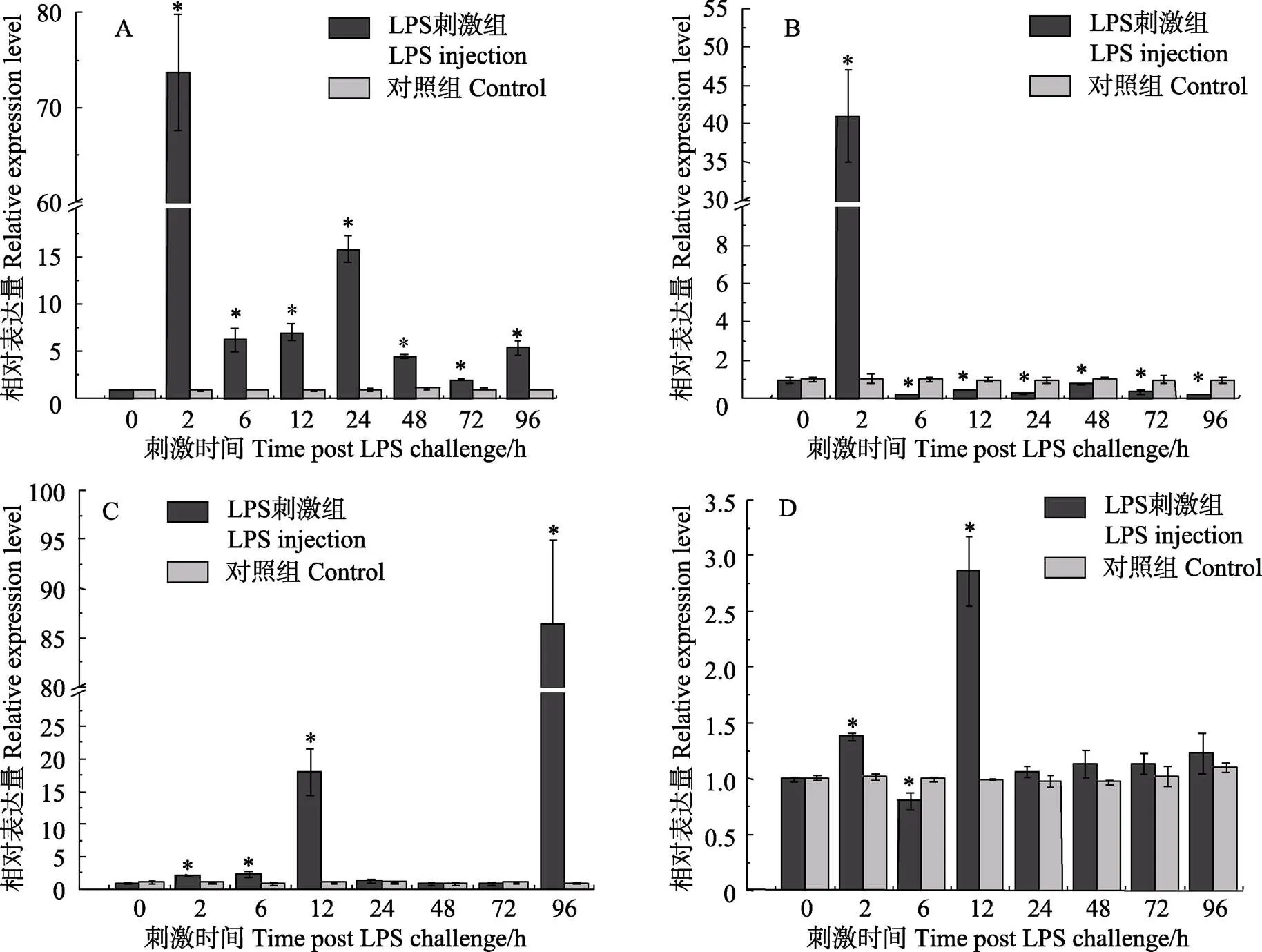
图6 LPS刺激后TfIL-15在不同组织的表达模式变化
A: 皮肤; B: 血; C: 肝脏 D: 脾. “*”表示实验组与对照组差异显著(<0.05)
A: Skin; B: Blood; C: Liver; D: Spleen. The asterisks indicate significant differences (<0.05)

图7 TfIL-15的原核表达
1: 诱导前细菌总蛋白; 2: 诱导后细菌总蛋白; 3: 超声破碎上清液; 4: 破碎后的包涵体; M: 蛋白Marker; 5: 纯化蛋白
Lane 1: Lysate ofwithout induction; Lane 2: Lysate ofwith induced with IPTG; Lane 3: Supernatant of recombinant pET-30a(+)TfIL-15 after ultrasonication; Lane 4: Sedimentation of recombinant pET-30a(+)/TfIL-15 after ultrasonication; M: Protein marker; 5: Purified protein
氨基酸序列多序列比对结果显示,不同物种动物IL-15的成熟肽区域的同源性较信号肽区域高,这可能由于成熟肽参与执行细胞因子的功能,而信号肽仅参与引导IL-15的胞内转运与分泌。在人类IL-15成熟肽序列中存在4个半胱氨酸残基,能形成两对二硫键(Bernard, 2004)。这4个半胱氨酸在鱼类lL-15同源蛋白序列中高度保守。蛋白质三级结构预测结果显示,TfIL-15与人类IL-15三级结构基本吻合,同属于4α螺旋家族,TfIL-15的2个二硫键位置与人类二硫键位置重合。这说明这些保守残基对于维持脊椎动物IL-15空间构象的形成和功能发挥可能起重要作用。三刺鱼、红鳍东方鲀、金头鲷和本研究松江鲈中,还含有另外的2个半胱氨酸,在三维结构预测中,这2个半胱氨酸形成了第3个二硫键(图3A)。这2个半胱氨酸是否确实形成新的二硫键,对于维持构象起到怎样的作用,尚需要进一步验证。
qRT-PCR结果显示,TfIL-15广泛表达于松江鲈各个组织器官中,这与以往研究鱼类IL-15的表达模式相同,并且类似于人类IL-15的广泛分布特征,这表明IL-15可能在多个生理过程中发挥重要的作用。值得注意的是,在卵中,IL-15也表现出了较高的转录水平表达。在对红鳍东方鲀的研究中也发现,IL-15在性腺中表达量较高。Das等(2015)在对露斯塔野鲮IL-15的研究中发现,IL-15在卵巢组织和未受精卵中也呈现出较高的表达量,而且在受精后发育初期表达量持续上调,都表明了IL-15转录本以母系遗传的形式给予待发育的后代被动免疫保护。IL-15在卵中遗传性的组成型高表达在卵排出体外、受精卵或幼体尚未获得自身免疫的时候,对于鱼类的体外受精及幼体发育过程中对抗水体复杂的病原微生物侵袭起着至关重要的作用。
为了进一步探讨TfIL-15在机体免疫中的作用,对LPS刺激后TfIL-15在松江鲈主要免疫组织内的表达情况进行了检测。结果发现,在血液和皮肤中,LPS刺激后2 h,TfIL-15迅速上调至最高峰,在肝脏和脾中12 h达到表达高峰,比皮肤和血液稍有延迟。这可能是因为血液作为承载各种免疫细胞和细胞因子的全身免疫器官,皮肤作为机体接触外界复杂水体环境的首个免疫组织,是最直接和快速的LPS应答和反应器官;而脾脏是鱼类红细胞、中性粒细胞和粒性白细胞等各种免疫细胞生产、贮存和成熟的主要场所,肝脏是鱼类多种重要免疫相关蛋白的合成场所,因而上调时间与皮肤和血液相比有所滞后。肝脏TfIL-15MRNA在刺激后96 h又出现一个新的高峰,这在对转铁蛋白、C-型凝集素、白介素1β等的研究中也有发现(Liu, 2012a、b; Yu, 2013),原因可能是首次免疫激活的免疫细胞产生的新的信号分子,又激活了新的免疫细胞类型,从而再次上调表达TfIL-15基因。在对红鳍东方鲀的研究中发现,在LPS刺激对IL-15的表达没有明显的影响。而在黑青斑河豚的研究中,LPS可引起几乎所有组织IL-15表达量的上调。本结果与黑青斑河豚的研究类似,推测可能IL-15在不同鱼体内的功能存在一定的差异。鱼类作为低等脊椎动物,其适应性免疫机制相对不完善,因此在抵御外界病原体入侵时主要依靠先天免疫发挥作用。细胞因子IL-15的上调表达正是适应了机体所需,通过其激活淋巴细胞、促进免疫相关蛋白表达等作用,在机体对抗外来病原入侵过程中发挥重要的抵御作用。鉴于TfIL-15可能在鱼类抵抗外界刺激的免疫反应中的作用,本研究成功地对松江鲈IL-15的成熟肽进行了体外表达,这为进一步研究TfIL-15蛋白的功能奠定了基础。
Bamford RN, DeFilippis AP, Azimi N,.The 5¢-untranslated region, signal peptide, and the coding sequence of the carboxyl terminus of IL-15 participate in its multifaceted translational control. Journal of Immunology, 1998, 160(9): 4418–4426
Bamford RN, Battiata AP, Waldmann TA. IL-15: The role of translational regulation in their expression. Journal of Leukocyte Biology, 1996, 59(4): 476–480
Bei JX. The homologues of interleukin (IL)-15 and IL-15 receptor subunits in Japanese puffeffish (): Cloning, identification and functional characterization. Doctoral Dissertation of Sun Yat-Sen University, 2006 [贝锦新. 红鳍东方鲀白细胞介素(IL)-15及其受体亚基同源基因的克隆鉴定和功能研究. 中山大学博士研究生学位论文, 2006]
Bei JX, Suetake H, Araki K,. Two interleukin (IL)-15 homologues in fish from two distinct origins. Molecular Immunology, 2006, 43(7): 860–869
Bernard J, Harb C, Mortier E,. Identification of an interleukin-15 alpha receptor-binding site on human interleukin-15. Journal of Biological Chemistry, 2004, 279(23): 24313–24322
Carson WE, Ross ME, Baiocchi RA,. Endogenous production of interleukin 15 by activated human monocytes is critical for optimal production of interferon-γ by natural killer cells in vitro. Journal of Clinical Investigation, 1995, 96(6): 2578–2582
Chen XZ, Zhang L, Yu SS,. Molecular cloning and expression analysis of ubiquitin-conjugating enzyme E2-D2 in roughskin sculpin,Acta Oceanologica Sinica, 2016, 37(10): 133–140 [陈学昭, 张雷, 于珊珊, 等. 松江鲈()泛素结合酶E2-D2基因的分子克隆及组织表达分析. 海洋学报, 2016, 37(10): 133–140]
Das S, Mohapatra A, Kar B,.Molecular characterization of interleukin 15 mRNA from rohu,(Hamilton): Its prominent role during parasitic infection as indicated from infection studies. Fish and Shellfish Immunology, 2015, 43(1): 25–35
Fang W, Xiang LX, Shao JZ,.Identification and characterization of an interleukin-15 homologue from. Comparative Biochemistry and Physiology Part B Biochemistry and Molecular Biology, 2006, 143: 335–343
Fang W. Molecular cloning, evolution and functional analyses of IL-15 subfamily and IL-15 receptor alpha chain family in teleost fish. Doctoral Dissertation of Zhejiang University, 2009 [方玮. 鱼类IL-15及其受体α链基因家族的分子克隆、进化和功能研究. 浙江大学博士研究生学位论文, 2009]
Fehniger TA, Caligiuri MA. Interleukin 15: Biology and relevance to human disease. Blood, 2001, 97(1): 14–32
Gunimaladevi I, Savan R, Sato K,. Characterization of an interleukin-15 like (IL-15L) gene from zebrafish (). Fish and Shellfish Immunology, 2007, 22(4): 351– 362
Kurys G, Tagaya Y, Bamford R,.The long signal peptide isoform and its alternative processing direct the intracellular trafficking of interleukin-15. Journal of Biological Chemistry, 2000, 275(39): 30653-30659
Lee YB, Satoh J, Walker DG,. Interleukin-15 gene expression in human astrocytes and microglia in culture. Neuroreport, 1996, 7(5): 1062–1066
Liu YY, Yu SS, Chai YM,. Transferrin gene expression in response to LPS challenge and heavy metal exposure in roughskin sculpin (). Fish and Shellfish Immunology, 2012a, 32(1): 223–229
Liu YY, Yu SS, Chai YM,. Lipopolysaccharide-induced gene expression of interleukin-1 receptor-associated kinase 4 and interleukin-1β in roughskin sculpin (). Fish and Shellfish Immunology, 2012b, 33(4): 690–698
Lodolce JP, Burkett PR, Koka RM,. Regulation of lymphoid homeostasis by interleukin-15. Cytokine and Growth Factor Reviews, 2002, 13(6): 429–439
Ma HX, Wang L, Wang LQ,. The different expression patterns of the gene Arginase Ⅱ inand the correlation between its expression andinfection. Progress in Fishery Sciences, 2018, 39(3): 44–52 [马慧鑫, 王磊, 汪林庆, 等. 牙鲆精氨酸酶Ⅱ基因的克隆以及免疫应答表达分析. 渔业科学进展, 2018, 39(3): 44–52]
Pérez-Cordón G, Estensoro I, Benedito-Palos L,.Interleukin gene expression is strongly modulated at the local level in a fish-parasite model. Fish and Shellfish Immunology, 2014, 37(2): 201–208
Ratthé C, Girard D. Interleukin-15 enhances human neutrophil phagocytosis by a Syk-dependent mechanism: Importance of the IL-15 alpha chain. Journal of Leukocyte Biology, 2004, 76(1): 162–168
Shi XL, Meng XH, Kong J,. cDNA cloning of the FBA gene inand its expression and functional analysis after WSSV infection. Progress in Fishery Sciences, 2018, 39(2): 112–119 [史晓丽, 孟宪红, 孔杰, 等. 中国明对虾基因克隆及其在白斑综合征病毒感染中的表达及功能分析. 渔业科学进展, 2018, 39(2): 112–119]
Sun SH, Ma P, Li H,. Prokaryotic expression and biological activity evaluation of recombinant IL15 from tiger pufferfish. Journal of Dalian Ocean University, 2015, 30(6): 598–603 [孙赛红, 马普, 李慧, 等. 红鳍东方鲀白介素15的原核表达及其活性测定. 大连海洋大学学报, 2015, 30(6): 598–603]
Wang DY, Huang JS, Guo MQ. Interleukin 15. Chemistry of Life, 1996, 16(3): 26–28 [王东勇, 黄建生, 郭明秋. 白介素15. 生命的化学, 1996, 16(3): 26–28]
Wang T, Holland JW, Carrington A,. Molecular and functional characterization of IL-15 in rainbow trout: A potent inducer of IFN-γ expression in spleen leukocytes. Journal of Immunology, 2007, 179(3): 1475–1488
Wang T, Zhang J, Li DJ,. Optimization on expression of recombinant human interleukin 21 and its biological activity analysis in vitro. Pharmaceutical Biotechnology, 2013, 20(4): 283–287 [王彤, 张娟, 厉道娟, 等. 重组人白介素21的原核表达与体外活性研究的优化. 药物生物技术, 2013, 20(4): 283–287]
Yu SQ, Wang SD. Characterization and breeding technology of roughskin sculpin,. Journal of Beijing Fisheries, 2008, 25(8): 50–52 [于诗群, 王世党. 松江鲈鱼的生物学特征及养殖技术.北京水产, 2008, 25(8): 50–52]
Yu SS, Yang H, Chai YM,. Molecular cloning and characterization of a C-type lectin in roughskin sculpin (). Fish and Shellfish Immunology, 2013, 34(2): 582–592
Characterization and Recombinant Protein Expression of Interleukin-15 from Roughskin Sculpin,
LIU Yingying, YU Shanshan, CHAI Yingmei, LIN Xiaopeng, ZHU Qian①
(School of Marine Science, Shandong University (Weihai),Weihai 264209)
Interleukin 15 (IL-15) is an important cytokine of fish immune system. In the present study, the IL-15 cDNA named as TfIL-15 was cloned from roughskin sculpin,. The full-length of TfIL-15 cDNA is 1140 bp, which contains a 5¢-UTR (untranslated region) of 165 bp, a 3¢-UTR of 453 bp and an open reading frame (ORF) of 522 bp, encoding a polypeptide of173 amino acids (aa) with a putative 59 aa-long signal peptide. Four out-of-frame AUG initiation codons, the negative translational regulators of mammalian IL-15 genes were also detected in the 5¢-UTR of TfIL-15. The protein sequence shared 23%~61% identity with reported fish IL-15s, displaying relatively high degree of variation. TfIL-15 homologues also contained four highly conserved cysteine residues allowing the formation of two disulfide bridges along with four predicted α-helices. Phylogenetic analysis grouped roughskin sculpin with other fish on a separated branch, excluded from mammalian and avian IL-15s. Quantitative real-time PCR (qRT-PCR) analysis showed that TfIL-15 was widely expressed in all detected tissues, with the highest expression in the heart.Post LPS challenge, TfIL-15 increased rapidly to the maximum of 74 folds and 41 folds compared with that of the control group at 2 h post challenge (hpc) in the skin and blood. The induction of TfIL-15 mRNA in the spleen and liver was 3 folds and 18 folds at 12 hpc. Interestingly, at 96 hpc, the expression of TfIL-15 in the liver was up-regulated again to the 86 folds higher than that of the control group. These results indicate that TfIL-15 may play an important role in fish innate immune response against microbial infections. Furthermore, the mature peptide of TfIL-15 was expressed inBL21 (DE3) cells successfully, laying a foundation for further research on the function of TfIL-15 protein.
Roughskin sculpin ();Interleukin-15 (IL-15); Gene clone; Relative expression; Recombinant protein expression
ZHU Qian, E-mail: qianzhu@sdu.edu.com
* 山东省自然科学基金(ZR2016CP10)和威海市科技局项目(1070413421706)共同资助[This work was supported by Natural Science Foundation of Shandong Province(ZR2016CP10), and Weihai Science and Technology Bureau (1070413421706)]. 刘莹莹,E-mail: liuying0631@163.com
祝 茜,教授,E-mail: qianzhu@sdu.edu.com
2018-06-25,
2018-07-31
Q78; S917
A
2095-9869(2019)04-0095-09
10.19663/j.issn2095-9869.20180625001
刘莹莹, 于珊珊, 柴迎梅, 林啸鹏, 祝茜. 松江鲈白介素15(TfIL-15)的结构特征与重组表达. 渔业科学进展, 2019, 40(4): 95–103
Liu YY, Yu SS, Chai YM, Lin XP, Zhu Q. Characterization and recombinant protein expression of Interleukin-15 from roughskin sculpin,. Progress in Fishery Sciences, 2019, 40(4): 95–103
(编辑 冯小花)
