产碳青霉烯酶肺炎克雷伯菌克隆复合群258的分子流行特征
2019-07-25刘婧娴阿旺穷吉
刘婧娴, 阿旺穷吉, 刘 瑛
作者单位:1. 上海交通大学医学院附属新华医院检验科,上海 200092;2. 西藏阜康医院妇产儿童分院检验科。
肺炎克雷伯菌是临床常见的条件致病菌之一,可引起血流感染、肺炎、尿路感染和中枢神经系统感染等。肺炎克雷伯菌遗传背景丰富多样,据巴斯德研究所统计,目前已检出3 000余种序列型(sequence type,ST)。近年,耐碳青霉烯类肺炎克雷伯菌(carbapenem-resistantKlebsiella pneumoniae,CRKP)的广泛播散已成为最棘手的全球性公共卫生问题之一,给人类健康带来了严重的威胁[1]。产碳青霉烯酶肺炎克雷伯菌(carbapenemase-producingKlebsiella pneumoniae,CPKP)是最常见的CRKP之一。目前在世界范围内流行的CPKP临床分离株以克隆复合群258(CC258)最为常见。本文旨在对CPKP的CC258流行特点作一综 述。
BURST是Feil等[2]设计和开发的用于计算亲缘关系的一种算法,它可以通过简单的计算方法,构建合理的细菌进化模型,并推演出原始基因型(founding genotype),该基因型细菌在进化过程中会衍生出很多非常相近的基因型,这些从一个原始基因型进化而来、遗传关系密切的基因型统称为克隆复合群。采用BURST算法对肺炎克雷伯菌的多位点序列分型(MLST)数据进行分析,可以发现CC258以ST258型为原始基因型,总共包含四十余种ST型。通过PubMed检索,CC258中,CPKP检出的ST型主要包括ST258、ST11、ST512、ST340、ST437和ST833等类型。
1 CPKP CC258流行特点
ST258型肺炎克雷伯菌是一个克隆杂交体,其80%基因组来自ST11型,其余20%来自ST442型肺炎克雷伯菌[3]。从2009年开始,有关ST258型CPKP检出或流行的报道逐渐增多。最初,ST258型CPKP在欧洲国家,如挪威、瑞典、波兰零星检出,随后其流行范围迅速扩张,在北欧及地中海以北的国家陆续检出,目前在美国、意大利、阿根廷、希腊、巴西、芬兰、波兰、德国等地均出现过暴发流行[4-7]。最新研究数据显示,ST258型肺炎克雷伯菌持续在北欧、南美洲及北美洲等地占据优势地位,如希腊、意大利、美国、巴西、阿根廷等地流行甚广,其次为芬兰、土耳其、匈牙利等[8-10]。我国ST258型CPKP检出相对较少,目前仅在江西省南昌市、香港地区、台湾地区分离到少数ST258型CPKP临床分离株[11-13]。
与ST258密切相关的序列型是ST11,该序列型与ST258相比只有一个管家基因(tonB)存在差异,同属CC258,两者亲缘关系较近。最初,在匈牙利、韩国、捷克等国家曾出现过产超广谱β内酰胺酶(ESBL)或AmpC酶的ST11型肺炎克雷伯菌流行[14-16]。2011年,Qi等[17]对中国五个省市的CRKP菌株进行分析,发现我国流行的CPKP以ST11型为主。此后,有关该克隆株CPKP的报道迅速增加,但是不同国家和地区检出的ST11克隆株所携带的碳青霉烯酶基因有所差异[18-19]。目前该克隆株在欧洲、亚洲、非洲、美洲均存在播散,其流行范围几乎遍布全球,中国是其流行“重灾区”,在沿海及中部地区省市流行较为广泛,目前黑龙江、吉林、辽宁、北京、河北、河南、安徽、江苏、上海、浙江、湖北、湖南、江西、四川、重庆、福建、广东、广西、云南、香港、台湾等省市均报道有ST11型CPKP检出[20-22]。
ST512与ST258克隆株相比,仅管家基因gapA存在差异。早在2006年,耶路撒冷地区曾检出ST512型CPKP[23]。2013年开始,该克隆株在意大利暴发流行的报道逐渐增多,其流行范围也进一步扩张至意大利周边国家如捷克、西班牙、德国等[24-27]。近年,在加拿大、澳大利亚、阿根廷、哥伦比亚、比利时、波兰等国家以及中国台湾地区也相继检出该克隆株[28-33]。
在CC258中,ST340是一种新兴的克隆类型。虽然ST340型 CPKP迄今为止尚未出现过大规模暴发流行,但是其与NDM基因在肠杆菌科细菌中的传播可能存在密切联系,因此也受到一定的关注。该克隆株的主要流行区域集中在亚洲地区,目前在泰国、印度、阿曼、韩国、中国、斯里兰卡、也门等国家均有检出,其中泰国流行情况较为严重[34-39]。此外,亦有个别欧美国家曾检出过ST340型CPKP,如希腊、加拿大、巴西、秘鲁、西班牙等[6,40-41]。Peirano等[40]报道,最初在加拿大发现的ST340型CPKP感染患者有印度旅行及就医史,因此推测这些菌株可能来源于印度。
ST437型CPKP的流行范围相对较为局限。Seki等[42]通过对2006-2009年巴西五个地区分离到的CRKP进行调查研究发现ST437型为主要流行株,之后的研究报道也显示该克隆株主要在巴西流行播散。此外,该克隆株在希腊曾出现过短暂的流行,在阿曼、加拿大、塞尔维亚、西班牙、斯里兰卡和中国也曾有零星检出[43-45]。
目前,仅意大利、委内瑞拉和以色列等国家有ST833型CPKP检出的相关报道[46-47]。一项调查研究显示,2014年4-7月委内瑞拉某医院共检出19株CPKP,所有菌株均为ST833型[47]。随后,在意大利里雅斯特医院1例来自委内瑞拉的儿童患者血培养标本中检出ST833型CPKP,提示该菌株可能来源于委内瑞拉。但是,之后再无该克隆株暴发流行的相关报道。
2 CPKP CC258产碳青霉烯酶情况
促使CPKP型CC258在全球范围内迅速播散的内在原因一直是研究者感兴趣的问题之一。Chmelnitsky等[48]发现CC258携带多种与细菌的运动、分泌、DNA重组和修复有关的独特基因,使该克隆株在进化过程中具有明显的优势,同时具有广泛传播的能力。近年,也有学者提出,ST11型等广泛流行的CPKP中存在干扰限制修饰(restriction and modi fication,RM)系统和毒素-抗毒素系统等,从而有助于耐药质粒的传播[49-50]。而耐药基因高效水平转移是公认的促进CPKP克隆株广泛播散的重要原因。综合文献报道发现,CPKP CC258对碳青霉烯类耐药的机制以产碳青霉烯酶为主,不同ST型克隆株所携带的耐药基因类型以及与水平转移有关的转座元件亦有所不同。
虽然ST258型CPKP流行范围非常之广,但是其所携带的碳青霉烯酶基因类型非常有限。2008年Hidalgo-Grass等[51]曾从1例耶路撒冷汉德森-希伯来大学医学中心住院患者肛门拭子标本中分离到携带KPC-9基因的ST258型CPKP,KPC-9基因位于Tn4401a转座子上,其所在的质粒与pKpQIL并无明显差异。Castanheira等[52]曾从1例接受造血干细胞移植患者的肾造瘘口引流的尿液标本中分离到1株同时携带KPC-2和VIM-4基因的ST258型CPKP。该菌株的KPC基因和VIM基因位于两个完全不同的质粒上,KPC基因被嵌入一个上游缺失99 bp序列的Tn4401a 转座元件,位于一个大小约120 kb的IncA / C质粒上,而VIM基因则位于一个大小约190 kb的IncFII质粒上,其上游存在整合子IntI1。除此之外,目前分离到的ST258型CPKP绝大多数均携带KPC-2或KPC-3基因。Peirano等[53]对全球ST258型肺炎克雷伯菌的菌株进行全基因组测序,发现该克隆株有2个不同的遗传片段(I和II),该克隆株主要与携带blaKPC密切相关,其中ST258-I携带blaKPC-2,ST258-II携带blaKPC-3。综合文献报道,ST258型CPKP所携带的KPC基因多数位于Tn4401a转座子上,其次为Tn4401以及Tn4401b。
ST11型CPKP所携带的碳青霉烯酶基因种类多种多样,包括A类、B类以及D类等十余种碳青霉烯酶基因(见表1),其中以KPC-2最为常见,OXA-48和NDM-1次之。此外,同一株ST11型CPKP同时携带多种碳青霉烯酶基因的情况也不少见[54-55]。该类型菌株所携带的KPC-2基因也绝大部分位于Tn4401a转座子上。韩国、巴西等地检出的ST11型CPKP,其KPC-2基因周围环境相对较多样化,部分KPC-2基因也存在于Tn4401b或Tn4401c上[56-57]。中国检出的ST11型CPKP绝大多数均携带KPC-2,但是KPC-2基因的周围环境多数以Tn1721为基础[58]。ST11型CPKP中的NDM-1基因多位于IncF质粒上,其播散可能与插入序列ISAba125有关[59]。该类型菌株所携带的OXA-48基因则多数位于IncL/M质粒上,OXA-48基因的周围环境则多以Tn1999为基础。突尼斯曾分离到1株携带OXA-204的ST11型肺炎克雷伯菌,该菌的OXA-204基因位于一个类似Tn 2016的转座子上。携带VIM基因的ST11型CPKP多分布在欧洲国家,如西班牙、捷克、匈牙利等[60]。而携带IMP基因的ST11型CPKP则多在中国台湾地区检出[61]。
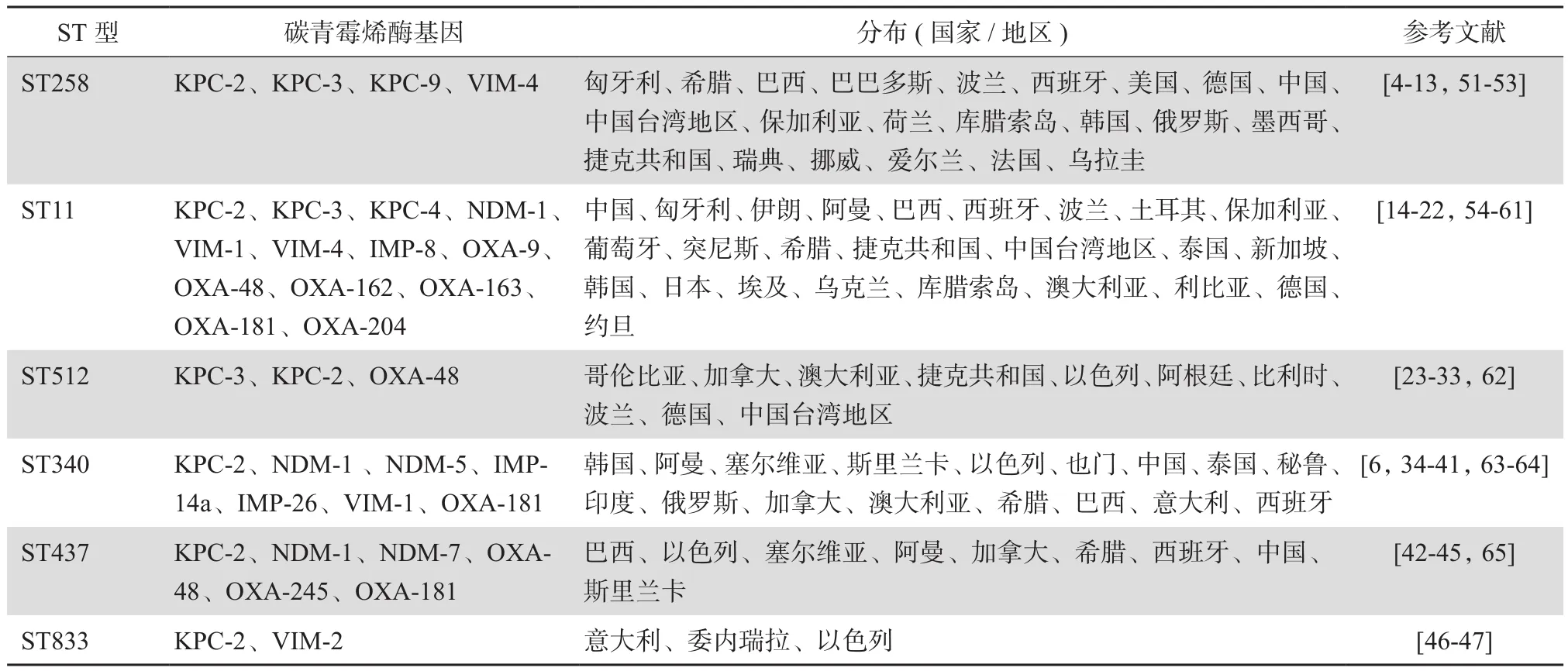
表1 CC258 CPKP所携带的碳青霉烯酶基因类型及其分布情况
根据目前报道,绝大多数ST512型CPKP携带KPC-3基因,KPC-3基因多位于FIIk质粒上,基因周围环境也以Tn4401a为主。目前仅在意大利及哥伦比亚地区检出过少量携带KPC-2基因的ST512型CPKP。意大利热那亚加列拉医院的一项调查研究显示,该医院最主要的CPKP流行株为ST512型,其中98%菌株携带KPC-3基因,仅2%携带KPC-2基因[62]。Ocampo等[31]对2012年6月-2014年6月哥伦比亚5所医院分离到的CRKP进行研究,仅发现1株携带KPC-2基因的ST512型菌株。
ST340型CPKP的流行规模不大,但是该菌株所携带的碳青霉烯酶基因却非常丰富。目前从该克隆株检测到的基因类型包括KPC-2、NDM-1、NDM-5、IMP-14a、IMP-26、VIM-1、OXA-181等,其中以NDM-1最为多见,其次为KPC-2(见表1)。韩国、也门、加拿大、俄罗斯、斯里兰卡等地均曾分离到产NDM-1酶的ST340型CPKP,而希腊、巴西、中国、秘鲁等地分离的ST340型CPKP则以携带KPC-2基因为主。虽然ST340型CRKP在泰国检出率较高,但菌株的耐药机制多为产ESBL或AmpC酶合并外膜蛋白表达异常,仅少数菌株产IMP-14a酶[39]。目前IMP-26阳性的ST340型CPKP仅在澳大利亚检出,NDM-5阳性的菌株则仅在西班牙有报道[63-64]。
目前从ST437型CPKP中检出的碳青霉烯酶基因包括KPC-2、NDM-1、NDM-7、OXA-48和OXA-245等。巴西、中国、以色列、希腊检出的该类型菌株均产KPC-2酶,KPC-2基因多位于Tn4401 b转座子上。也门和加拿大等地曾检出产NDM-1酶菌株,西班牙曾出现过产NDM-7酶菌株的小规模流行。Oteo等[65]发现1株分离自西班牙马拉加地区的ST437型CPKP携带OXA-245基因。Novović等[43]在塞尔维亚分离到1株耐多黏菌素且产OXA-48酶的ST437型CPKP。Zhu等[45]最近在斯里兰卡一所教学医院分离到2株携带OXA-181基因的ST437型CPKP。
委内瑞拉所检出的ST833型CPKP均同时携带KPC-2和VIM-2基因,其中KPC-2基因位于Tn4401b转座子上[47]。意大利和以色列分离到的ST833型CPKP均仅携带KPC-2基因,前者的KPC基因位于ColE1质粒上,后者则位于100 kb左右含FIIk和FIBk复制子的质粒上[46]。
3 总结
CPKP在全球范围内广泛流行归因于CC258的成功传播,尤其是ST258型和ST11型菌株。ST258克隆株的遗传背景相对较为保守,在流行过程中进化相对较为缓慢。而ST11型克隆株则像一个耐药基因的储存库,在流行范围不断扩张的同时,其携带的耐药基因种类也呈现明显的多样化特点,值得注意的是,我国流行的CPKP绝大多数为该克隆类型,因此,临床检出此类菌株时,应引起足够重视,加强医院感染控制措施和监测工作。虽然,目前ST512、ST340以及ST437等类型尚未引起大规模的暴发流行,但是其检出范围有不断扩大的趋势,也应引起足够重视。
