福建三明格氏栲和杉木叶片养分季节动态及再吸收特征
2019-07-24孟庆权葛露露杨馨邈何宗明胡欢甜邱岭军
孟庆权,葛露露,王 俊,杨馨邈,林 宇,何宗明* ,胡欢甜,邱岭军
(1.福建农林大学林学院/国家林业和草原局杉木工程技术研究中心,福州 350002;2.福建省长乐大鹤国有防护林场,福州 350212)
植物叶片养分再吸收是指植物组织对衰老叶片中的养分进行重复利用的过程[1-3],使植物生长减少了对根系养分吸收的依赖[4],是植物体对贫瘠环境的一种适应策略[5],也是植物保存养分、增强竞争力、提高生产力的重要策略[6-7]。养分再吸收在植物叶片衰老过程中普遍存在,它可以减少凋落物分解产生的养分损失,增加养分的利用效率,叶片的养分可以被重新利用,这对于满足新生组织在树体中的营养需求是十分必要的[8]。
从20 世纪20年代开始,国外就有关于养分再吸收过程及其影响因子的研究,到目前仍有大量养分再吸收的研究报道[9-10],直到20 世纪90年代末国内才有了关于养分再吸收的研究,而国内关于养分再吸收的研究主要集中在樟子松(Pinus sylvestris)、短枝木麻黄(Casuarina equisetifolia)、赤桉(Eucalyptus camaldulensis)、杉木(Cunninghamia lanceolata)和马尾松(Pinus massoniana)等树种[11-15],且多数偏重于 N、P 元素的研究,除 N、P 以外,K、Ca、Mg 等金属元素也是植物生长过程不可或缺的大量营养元素[16],K 主要作用于碳水化合物的合成和转移,缺K 会导致植物光合作用减弱[17],Ca 和Mg 是植物叶片中多种酶的催化剂,能有效促进植物叶片细胞的有丝分裂和蛋白质合成[18]。
在我国亚热带地区,由于人类活动的长期干扰,原生的常绿阔叶林所剩无几,取而代之的是大面积的次生林和人工林等。杉木(Cunninghamia lanceolata)是重要的速生用材树种,具有生长快、再生力强、经济价值高、栽培地区广等优点,在林业生产上占据了重要的地位。格氏栲(Castanopsis kawakamii)属国家二级保护植物,是壳斗科常绿阔叶大乔木、亚热带珍稀濒危植物之一。本文以20 世纪60年代末格氏栲天然林皆伐后种植的格氏栲和杉木人工林为研究对象,试图从叶片衰老过程中N、P、K、Ca和Mg 含量及其再吸收率的季节变化来探讨人工林对贫瘠生境的适应和竞争机制,从而为人工林的养分管理和合理经营提供科学的理论依据。
1 材料和方法
1.1 研究区概况
本研究区位于福建省三明市莘口镇福建农林大学莘口教学林场,地理坐标(117°27'11″~117°29'45″E,26°9'30″~26°12'49″N,海拔 250~604.2 m),属武夷山脉东伸支脉,为中亚热带季风性气候,受热带海洋气团和大陆气团交替控制,气候温暖湿润,年均气温为19.1 ℃,无霜期300 d,年均降雨量为1 750 mm,年均相对湿度81%,土壤类型主要为沙质页岩发育的山地红壤,土层厚度>80 cm。
格氏栲人工林层次结构简单,乔木层树种单一(格氏栲),灌木层主要有山血丹(Ardisia punctata)、黄绒润楠(Machilus grijsii)、山矾(Symplocos sumuntia)和毛冬青(Ilex pubescens)等,草本层种类较少,主要有狗脊(Woodwardia japonica)、山姜(Alpinia japonica)、黑莎草(Gahnia tristis)、扇叶铁线蕨(Adiantum flabellulatum)。
杉木人工林灌木层主要有栲树(Castanopsis fargesii)、鸭脚茶(Bredia sinensis)、虎皮楠(Daphniphyllum oldhami)和毛冬青等,草本层主要有狗脊、山姜、金星蕨(Parathelypteris glanduligera)。
在研究区内选取相同林龄(50 a),代表性强的格氏栲人工林和杉木人工林样地,在样地内分别随机设置 3 个 25 m×25 m 的样方,共 6 个,林分基本情况见表1。
1.2 实验设计与采样
于 2017年 3、6、9、12月底,在三明格氏栲人工林和杉木人工林的6 个样方内对角线选取5 株生长良好的格氏栲(杉木)作为标准木,考虑到植株不同高度的叶片养分含量可能差异较大,采用人工攀爬的方式在标准木上、中、下部分别摘取6~8 枝,并选取完全展开、没有病虫害或斑点的成熟叶30 枚,将同一样方内5 株标准木上采集的成熟叶样品混合;衰老叶的收集主要通过采摘树上已经枯黄且没有凋落的叶片,衰老叶片为黄色很容易区别于成熟叶,将同一样方内5 株标准木上采集的衰老叶样品混合,带回实验室待测。先在105 ℃烘箱内杀青,然后设定70 ℃烘干至恒重,用高速万能粉碎机研磨后过100 目筛放入自封袋密闭保存,进行养分测定。
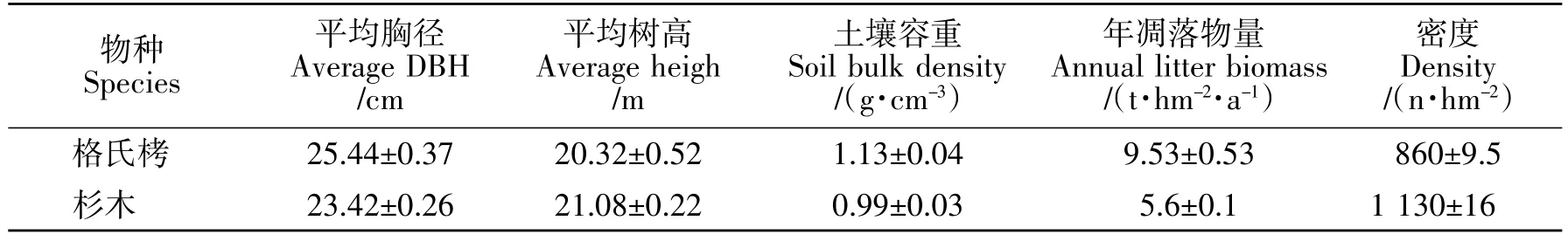
表1 试验林林分基本情况Table 1 Basic starts of the institutional forest
1.3 叶片养分含量测定
叶片N 含量采用全自动碳氮分析仪(Elementar ELVario Max,德国)直接测定。取一定研磨好的样品用浓硫酸-高氯酸消煮法制备成待测液后用钼锑钪比色法测定P 含量,K、Ca、Mg 含量均采用原子吸收分光光度法测定[19]。
1.4 养分再吸收的表示
养分再吸收通常用养分转移度(nutrient resorption proficiency)和养分再吸收率(nutrient resorption efficiency)表示,前者是指凋落叶中养分含量相对转移极限值的实现程度,通常用衰老叶中养分浓度表示。养分再吸收率=(成熟叶养分浓度-衰老叶养分浓度)/成熟叶养分浓度×100%。
1.5 数据分析
采用Microsoft Excel 2003 进行数据处理及绘图,采用SPSS 17.0 统计分析软件对两种人工林成熟叶和衰老叶中不同季节养分含量和再吸收率进行单因素方差分析(one-way ANOVA)和多重比较分析(LSD),对两种人工林平均叶片养分含量及其再吸收率进行独立样本T 检验,并对成熟叶和衰老叶养分含量与再吸收率进行线性回归分析。
2 结果与分析
2.1 格氏栲和杉木成熟叶养分含量及其季节动态
从表2可见,格氏栲成熟叶 N、P、K、Ca、Mg 含量均值分别为 15.67、0.93、12.12、5.83、8.20 g/kg,变异系数分别为 14.70%、14.01%、17.44%、18.92%、17.85%,杉木成熟叶 N、P、K、Ca、Mg 含量均值分别为 13.67、1.00、12.93、4.41、7.45 g/kg,变异系数分别为 12.88%、15.05%、13.87%、17.71%、22.43%,格氏栲叶片 N、Ca、Mg 含量显著高于杉木(P<0.05),P 含量显著低于杉木,K 含量差异不显著,格氏栲叶片N、K、Ca 含量变异系数高于杉木,而 P、Mg 含量变异系数低于杉木。
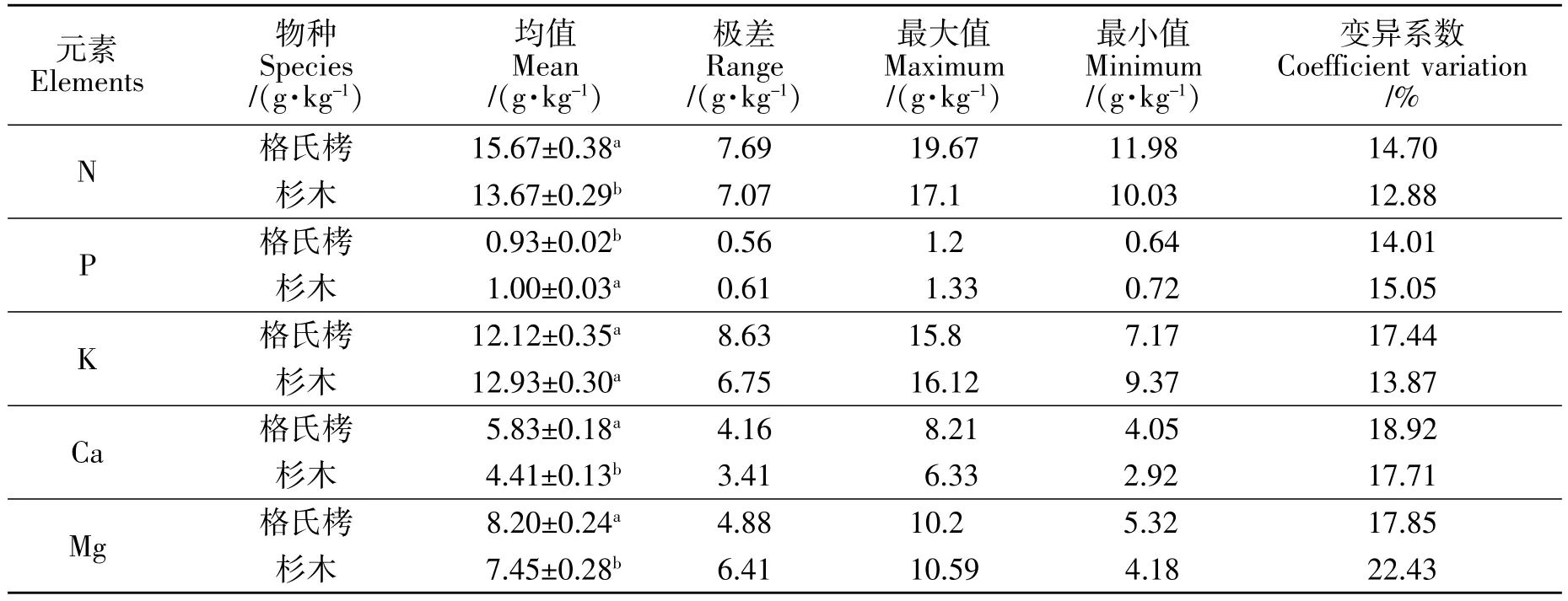
表2 格氏栲和杉木成熟叶养分含量及其变异系数Table 2 Nutrient concentrations and its coefficient of variation in mature leaves of C.kawakamii and C.lanceolata
由表3可知,两种人工林叶片 N、P、K、Ca、Mg含量的季节变化不一致,格氏栲叶片N 含量在各季节间均差异显著(P<0.05),随季节变化表现出先降低后增高的趋势,呈“V”字形,春夏季不断下降,秋季最低,然后迅速上升,冬季达到一年中的最高水平,而杉木叶片N 含量基本保持不变,各季节间无显著差异(P>0.05),格氏栲与杉木叶片N 含量在春冬季节差异显著;格氏栲叶片P 含量表现为先升高再降低的趋势,夏季达到最高,随后保持稳定,而杉木叶片P 含量表现为秋冬季显著高于春夏季,格氏栲与杉木叶片P 含量在春冬季节差异显著。格氏栲叶片K 含量为春季最低,显著低于其他季节,而杉木叶片K 含量为春季最高,显著高于其他季节,格氏栲与杉木叶片K 含量在春季差异显著;格氏栲和杉木叶片Ca 含量均为冬季最低,格氏栲与杉木叶片Ca 含量在不同季节均差异显著,不同季节格氏栲叶片Ca 含量均高于杉木;格氏栲和杉木叶片Mg 含量随季节变化均呈现出先降低后升高的趋势,秋季最低,格氏栲和杉木叶片Mg 含量在冬季差异显著。
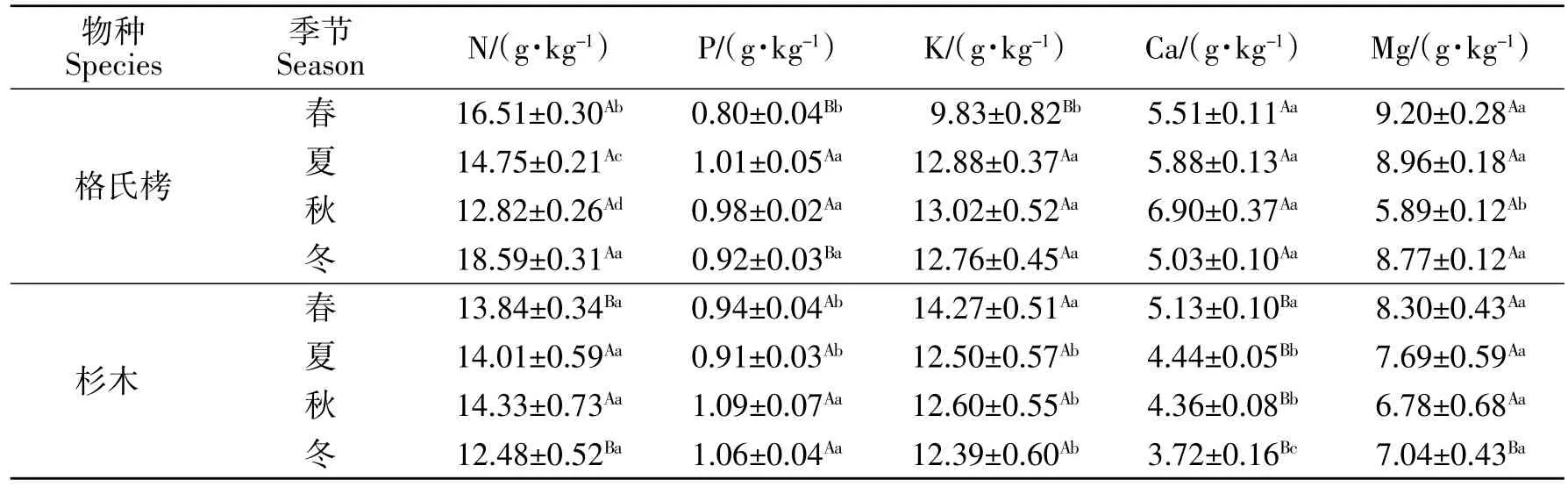
表3 格氏栲和杉木成熟叶养分浓度季节动态Table 3 Seasonal dynamics of nutrient concentrations in mature leaves of C.kawakamii and C.lanceolata
2.2 格氏栲和杉木叶片养分转移度及其季节动态
从表4可见,格氏栲和杉木衰老叶养分浓度在不同季节差异显著,衰老养分浓度越低表示其转移度越高,格氏栲叶片N 转移度为秋季最高春季最低,P 转移度为春季最高,显著高于其他季节,Mg 转移度为秋季最高,春夏冬季节差异不显著,K、Ca 转移度在不同季节间差异不显著;而杉木叶片P 转移度为春夏季显著高于秋冬季,而N、K、Ca、Mg 转移度在不同季节间差异不显著,不同树种间,杉木叶片N、K、Ca 转移度显著高于格氏栲,而P、Mg 转移度差异不显著。
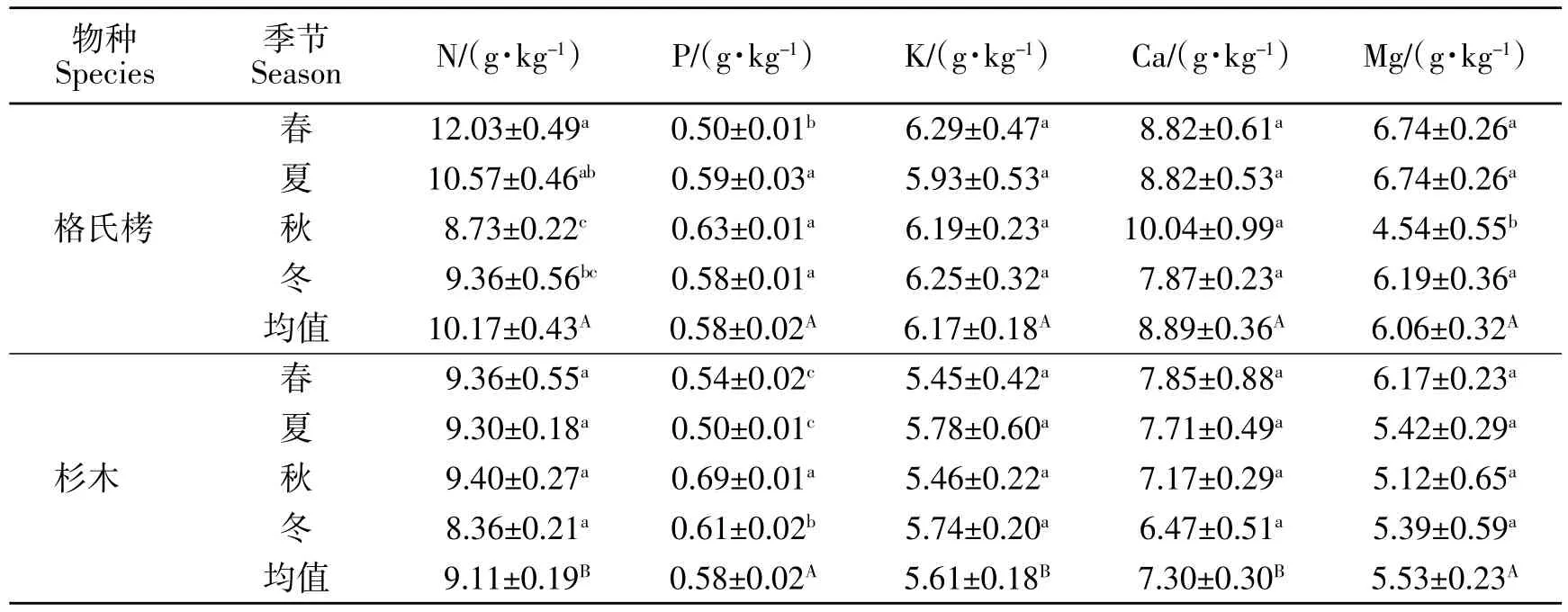
表4 格氏栲和杉木衰老叶养分浓度季节动态Table 4 Seasonal dynamics of nutrient concentrations in senescent leaves of C.kawakamii and C.lanceolata
2.3 格氏栲和杉木叶片养分再吸收率及其季节动态
从图1可见,杉木叶片养分再吸收率随季节的变化没有表现出显著差异(P >0.05),而格氏栲叶片N 再吸收率为冬季显著高于春夏秋季(P<0.05),而K 再吸收率为春季显著低于夏秋冬季,P、Ca、Mg 再吸收率在不同季节差异不显著。由表5可知,不同人工林之间,格氏栲 N、P、K、Ca、Mg 再吸收率分别为 34.21%、37.65%、48.27%、-53.09%、25.72%,杉木为 33.19%、41.35%、56.23%、-66.67%、24.87%,其中杉木叶片P、K 再吸收率显著高于格氏栲,N、Mg 再吸收率无显著差异,Ca 没有被再吸收。
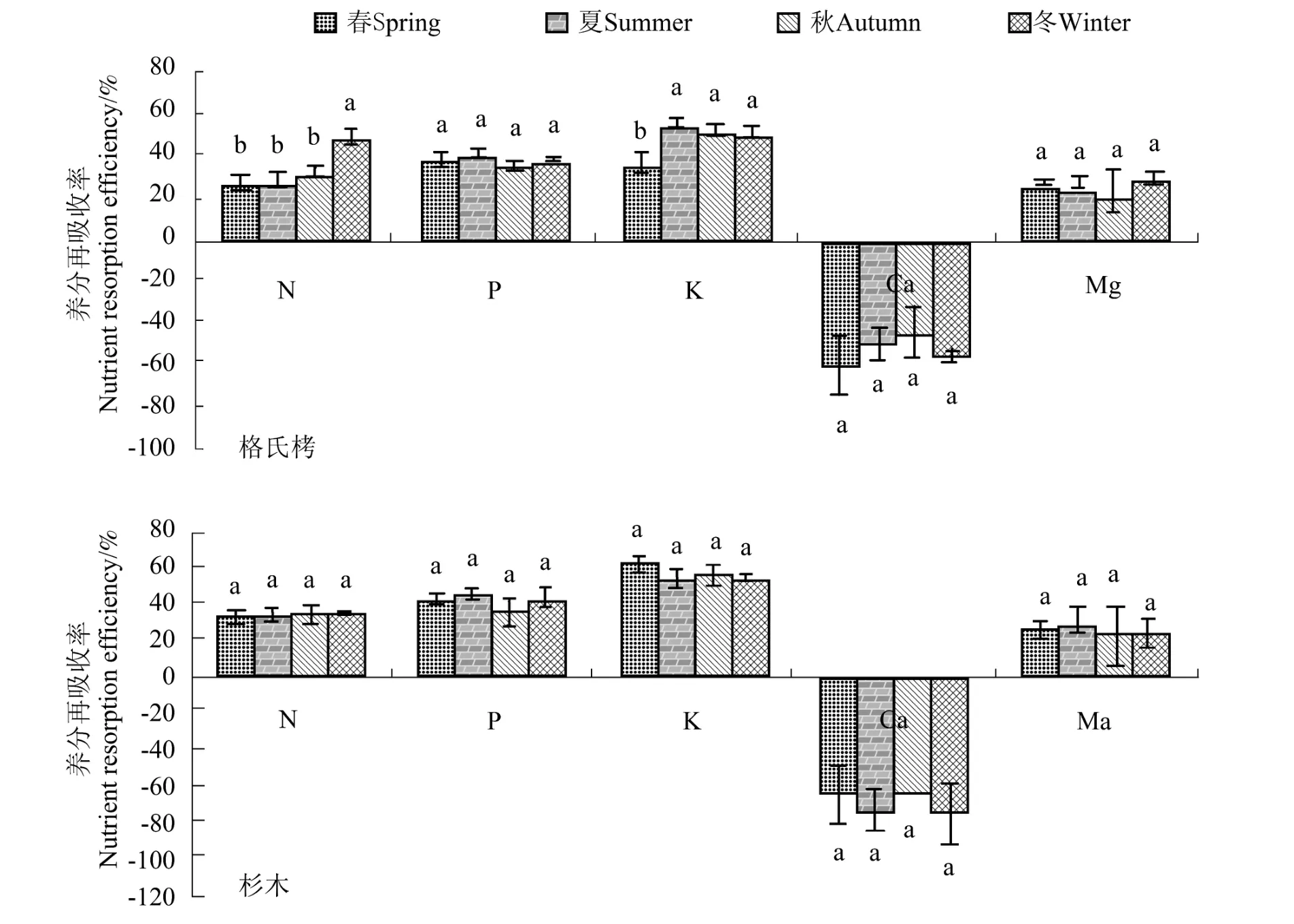
图1 叶片养分再吸收率季节动态Figure 1 Seasonal dynamics of leaf nutrient resorption rate
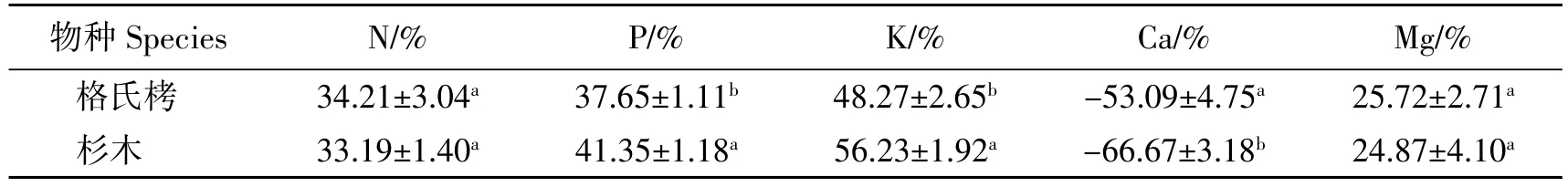
表5 不同人工林叶片养分再吸收率Table 5 Leaf nutrient resorption rate of different plantations
2.4 养分再吸收率与叶片养分含量相关性
以养分含量为横坐标(x)、其再吸收率为纵坐标(y),对成熟叶和衰老叶中各养分含量与再吸收率进行线性回归分析,结果见表6。
由表6可知,格氏栲和杉木成熟叶中K 含量与其再吸收率呈极显著的线性正相关(P<0.01),回归方程为y=4.655x-8.175(R2=0.645)和y=4.180x+2.160(R2=0.553),格氏栲成熟叶中 N 含量与 N 再吸收率呈显著的线性正相关(P<0.05),回归方程分别为 y=2.893x-11.114(R2=0.380)。
由表6可知,格氏栲和杉木衰老叶养分含量均与其再吸收率呈线性负相关(P<0.05),其中格氏栲衰老叶N、K 含量与其再吸收率呈显著的线性负相关,回归方程分别为 y=-4.433x+79.316(R2=0.386)、y=-8.618x+101.401(R2=0.334),杉木衰老叶 K、Ca含量与其再吸收率呈极显著的线性负相关(P<0.01),回归方程分别为 y=-8.627x+104.623(R2=0.386)、y=-12.630x+94.711(R2=0.520)。
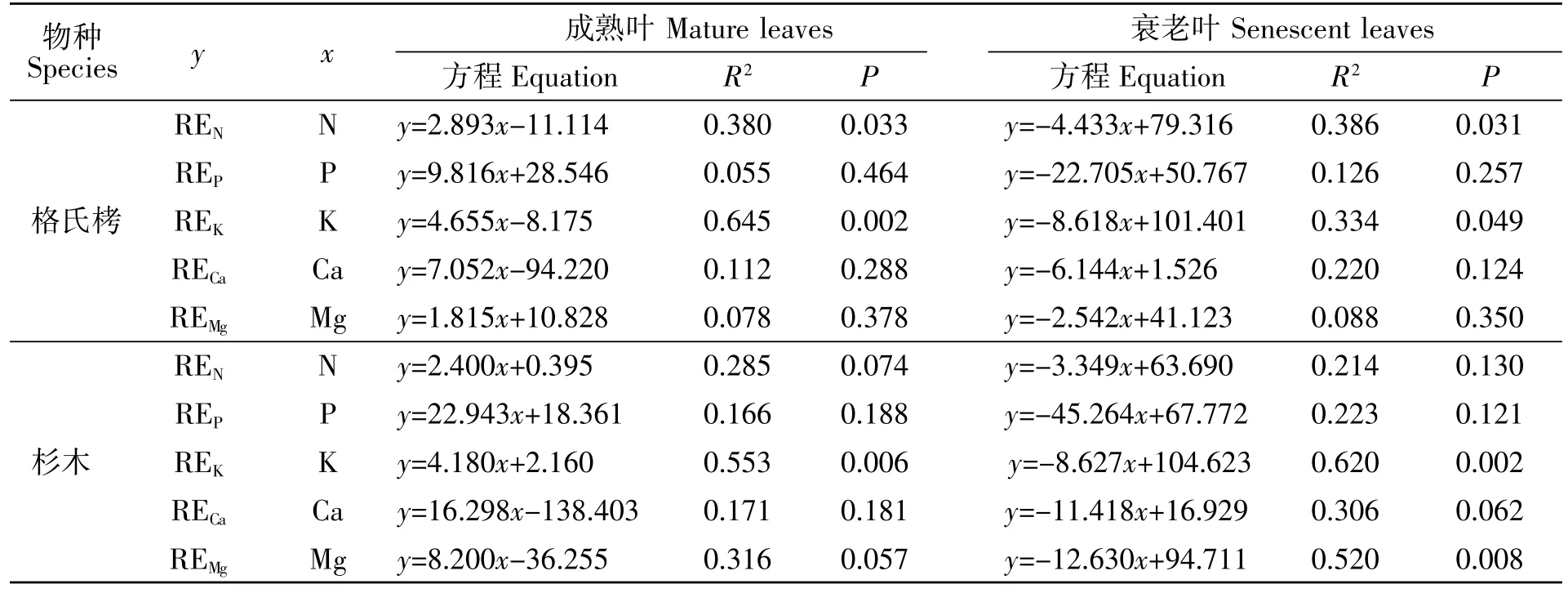
表6 叶片养分浓度与再吸收率的相关性(n=12)Table 6 Correlation between nutrient elements contents and their resorption rates
3 讨论
3.1 叶片养分和再吸收率季节动态
叶片是对树体养分盈余反应最敏感的器官,它既是地下运输来的矿质元素的贮存库,又是果实生长发育所需矿质元素的供给源。叶片养分含量的动态变化是植物遗传因素、物候期以及外界环境(如降水淋溶等)共同作用的结果[20]。由于各元素的生理机能不同,各元素含量也存在一定的差异,总体而言,两种人工林成熟叶养分含量大小顺序为N>K>Mg>Ca>P,而衰老叶养分含量大小顺序为 N>Ca>K>Mg>P,不同季节顺序略有不同。
养分再吸收在植物营养保存方面起着关键作用,可以减少养分流失,降低对环境的依赖性,是植物对贫瘠环境的一种适应[21-22]。两种人工林成熟叶到衰老叶阶段,Ca 含量逐渐增加,由于Ca 不具有流动性,不能被再吸收,而 N、P、K、Mg 含量呈下降趋势,说明N、P、K、Mg 元素在叶片衰老过程中发生了养分再吸收,两种人工林不同元素再吸收率大小顺序均为 REK>REP>REN>REMg。
本研究中格氏栲和杉木人工林叶片N 再吸收率分别为34.21%、33.19%,格氏栲成熟叶与衰老叶N 含量均高于杉木,但两者N 再吸收率无显著差异,可能是由于格氏栲对N 的需求量高造成的[23],也可能是因为叶片N 的来源不唯一,植物可以通过根系吸取土壤中的有效态N,另外大气氮沉降也是氮的重要来源[24],而两种人工林叶片P 的再吸收率差异显著,杉木(41.35%)显著高于格氏栲(37.65%),一方面可能是由于杉木人工林凋落物量少,养分归还土壤少,植物需要更高的再吸收率来适应贫瘠环境,另一方面杉木叶片延迟脱落,延长了再吸收时间。2 种人工林P 的再吸收率均大于N 的再吸收率,说明P 优先再吸收,这与福建武夷山[25]和浙江天童山[26]生态系统中限制性元素优先再吸收的结果一致。
两种人工林K 的再吸收率差异显著,杉木(56.23%)高于格氏栲(48.27%),叶片表面粗糙且具有较高养分含量的叶片可能会更易被降水淋溶[27],而K 属于极易淋溶的元素,杉木叶片表面较格氏栲粗糙,养分含量也高于格氏栲,导致杉木叶片淋溶损失了较多的K,最终表现为较高的K 再吸收率,因此计算的养分再吸收率也包括了部分的淋溶损失。
Mg 是叶绿素的组成成分,是光合作用必要的元素之一,叶片衰老前随叶绿素的水解而被释放[28],本研究结果中格氏栲Mg 的再吸收率为25.72%,杉木为24.87%,2 种人工林之间无显著差异,邹邦基等[29]研究表明Mg 元素的再吸收率是以植物体内的实际需求为准,幼叶若缺乏Mg 元素,就会发生很高的养分转移率,但对该研究区2 种人工林的研究显示Mg 再吸收率较低,低于干热河谷赤桉林[13]和长白山主要木本植物[30],说明该研究区内2 种人工林幼叶生长不缺Mg 元素。
不同于其他元素再吸收率,本研究2 种人工林叶片Ca 的再吸收率均为负值,与严思维等[13]、曾德慧等[11]的研究结果一致,呈累积状态,且表现为杉木人工林>格氏栲人工林,说明杉木人工林叶片凋落前Ca 的累积能力较强。但王文卿等[31]统计发现杉木Ca 元素再吸收率为正值,本研究结果中杉木人工林再吸收率为-66.67%,与林宝平等[14]对不同林龄杉木人工林Ca 的再吸收率接近,但与王文卿等[31]的统计结果(0.7%)差异较大,可能是由于衰老叶样品取样差异造成的,本研究采集的衰老叶为树枝上已经枯黄的叶片,而王文卿等[31]所统计的为凋落叶,杉木枝条存在宿存现象,叶片在枯死枝条经多年分解才能从树干脱落,期间Ca 大量流失[14]。
杉木人工林叶片P、K 再吸收率显著高于格氏栲人工林,说明杉木对养分的利用率相对更高,更能适应贫瘠生境。2 种人工林叶片养分含量随季节变化差异显著但不同季节叶片养分再吸收率差异不显著,R.Aerts 等[5]的研究表明,温度变化对养分再吸收率没有显著影响,叶功富等[7]研究也表明短枝木麻黄N 和 P 再吸收率在季节之间的差异不显著。因此我们推测养分再吸收是树种的特有属性,不随植物物候期的变化而改变。
3.2 叶片养分含量与再吸收率的相关性
成熟叶片的养分含量对养分再吸收效率的影响目前还没有形成统一定论。有学者认为,养分再吸收效率随成熟叶养分含量的增加而降低[32],也有的学者认为,较高的养分含量促进养分的再吸收效率[33]。本研究中格氏栲和杉木人工林成熟叶N、K 含量与养分再吸收率呈显著正相关,说明格氏栲和杉木成熟叶N、K 含量高可以促进N、K 元素的再吸收,而成熟叶中其他养分元素与养分再吸收率无显著相关关系,可能是因为随着养分有效性的变化,林木会优化其通过根系从土壤吸收养分的成本消耗与通过养分再吸收机制回收养分的成本消耗,进而对不同的养分元素表现出不同的利用策略[34],而格氏栲和杉木衰老叶养分含量均与其养分再吸收率呈负相关,这与严思维等[13]、林宝平等[14]的研究结果一致,说明高的养分再吸收率降低了衰老叶养分含量。
4 结论
格氏栲和杉木人工林成熟叶片中养分元素表现出明显的季节动态,N、Mg 在生长旺季最低,而P含量为生长旺季最高,Ca 含量为冬季最低。格氏栲和杉木叶片中N、P、K、Mg 都有一定程度的回收,其中杉木叶片的养分再吸收率更高,说明杉木更能适应贫瘠环境。格氏栲和杉木衰老叶片养分含量与其再吸收率均呈负相关,说明养分转移度越高,养分再吸收率越大。
本研究结果仅仅依据叶片衰老前后的养分浓度计算养分再吸收率,但叶片养分的最高浓度并不一定出现在叶片衰老前,如要真实反映叶的养分再吸收率,就必须选择叶生活史中养分含量最高的为计算依据,因此,需要采集不同叶龄的叶片进行养分含量的测定,以便更准确地计算养分再吸收率,为人工林的养分管理提供科学依据。
