长江口及邻近海域凤鲚4个不同地理群体的形态差异
2019-07-03苗中博
杨 琴,赵 峰,3,宋 超,张 涛,苗中博,庄 平
(1.中国水产科学研究院东海水产研究所,上海 200090;2.上海海洋大学水产与生命学院,上海 201306;3.农业农村部东海与长江口渔业资源环境科学观测实验站,上海 200090)
凤鲚(Coilia mystus),隶属于鲱形目(Clupeiformes),鳀 科 (Engraulidea),鲚 属(Coilia),俗称凤尾鱼、烤子鱼、籽鲚等,为河口洄游性鱼类,在我国东海、黄海和渤海的近海海域均有分布[1-2]。凤鲚曾是长江口“五大渔汛”之一,1974年捕捞产量高达5 284 t,占长江口渔业总捕捞量的 48.6%[3]。20年来,在栖息地环境变迁和过度捕捞等因素的影响下,凤鲚资源急剧衰退,到2011年长江口的捕捞产量仅为107.6 t[4],现今已几乎不能形成渔汛[3-5],捕捞价值基本丧失,农业农村部也通告要求“自2019年2月1日起禁止刀鱼、凤鲚、河蟹三种天然资源的生产性捕捞”,因此长江口凤鲚资源的养护管理工作亟待开展。
凤鲚属于短距离洄游型鱼类,平时栖于浅海,呈分散型生活。每年繁殖盛期为5—7月,大量成熟个体集群洄游至不同河口咸淡水或淡水区域进行生殖洄游,产卵结束后,亲鱼返回近海,仔幼鱼在河口觅食。不同河口区繁殖的凤鲚群体间个体存在迁入或迁出现象,群体结构复杂。对长江口凤鲚资源进行评估和管理,需要考虑到其与附近河口区凤鲚地理群体间的差异及联系,以避免管理策略偏差[6]。鱼类形态是基因和环境相互作用的产物,在鱼类种群鉴别及划分中起到重要作用,主要包括种间及种内的群体划分[7-8]。随着测量及统计方法的不断改进,采用传统形态测量及框架法与多元分析相结合的方法,除了已应用于华鳊属(Sinibrama)[9]、石爬鮡(Euchiloglanis sp.)[10]、鲈属(Perca)[11]和鲻科(Mugilidae)[12]等的种间差异性研究,也在刀鲚(Coilia nasus)[13]、红 鳍 东 方 鲀 (Takifugu rubripes)[14]、多 鳞 四 指 马 鲅 (Eleutheronema rhadinum)[15]、日本蟳(Charybdis japonica)[16]、罗氏 沼 虾 (Macrobrachium rosenbergii)[17]和 锦 鲤(Cyprinus carpio)[18]等水生生物的种内分化研究中得到应用。本研究采用传统形态测量法和框架法描述凤鲚形态的几何特征,再利用多元统计分析方法对长江口及其邻近海域4个地理群体凤鲚的形态差异进行综合分析,旨在查明长江口及其邻近海域不同凤鲚群体间的差异性及亲缘关系,为长江口凤鲚资源养护和管理提供科学支撑。
1 材料与方法
1.1 实验材料
本研究所用样本分别采集于江苏吕四(121°35′51″E、32°5′49″N)、上海崇明(121°44′57″E、31°23′22″N)、浙江舟山(122°2′27″E、29°36′28″N)和浙江温州(120°57′20″E、28°5′20″N)4个近岸海域(图1),各40尾成鱼,冷冻保存,实验室测量、称重及拍照。采集时间为2018年9—10月,凤鲚繁殖期基本结束,不存在怀卵的雌性凤鲚,因此无法从外形上区分样本性别,故选用雌雄混合样本用于形态分析(表1)。
1.2 形态参数测定
利用传统形态测量法和形态框架法对4个海域的凤鲚样本进行测量。参照解剖学坐标点以及测量指标,采用Image Pro Plus 6.0软件进行测量。其中,传统形态学性状测量包括11个参数(图2),即全长(FL)、体长(BL)、体高(BD)、头高(HD)、头长(HL)、眼后头长(HBE)、躯干长(STL)、尾长(TL)、吻长(SL)、眼径(ED)、尾柄高(CPH);框架测量指标19个,即D1~D19(图3)。
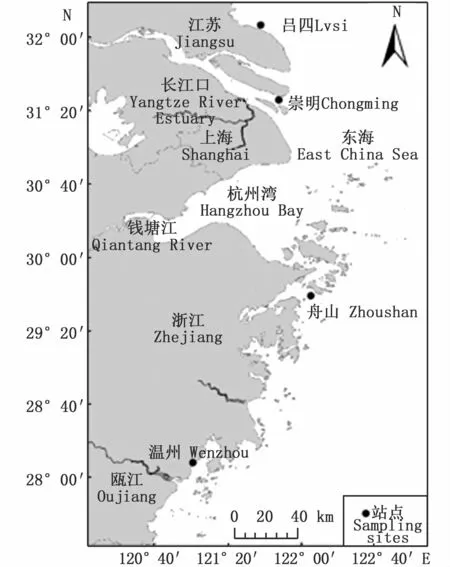
图1 采样地点Fig.1 Sampling sites
1.3 数据分析
利用Microsoft Excel初步统计所有样本11个传统形态学特征数据及19个形态框架数据,为消除样本个体大小差异对形态特征的影响,将以上除体长(BL)外29个形态数据与其体长(BL)的比值作为形态度量分析的性状值,从而得到29个形态比例性状参数,然后采用SPSS 23.0统计软件对4个群体凤鲚的29个形态比例性状参数进行非参数检验、主成分分析、判别分析和聚类分析,通过分析结果探讨群体间的差异性及关联性。

表1 凤鲚样本信息Tab.1 Sampling information of C.mystus

图2 凤鲚形态学测量位点Fig.2 Morphological and mark points for measurement of C.mystus
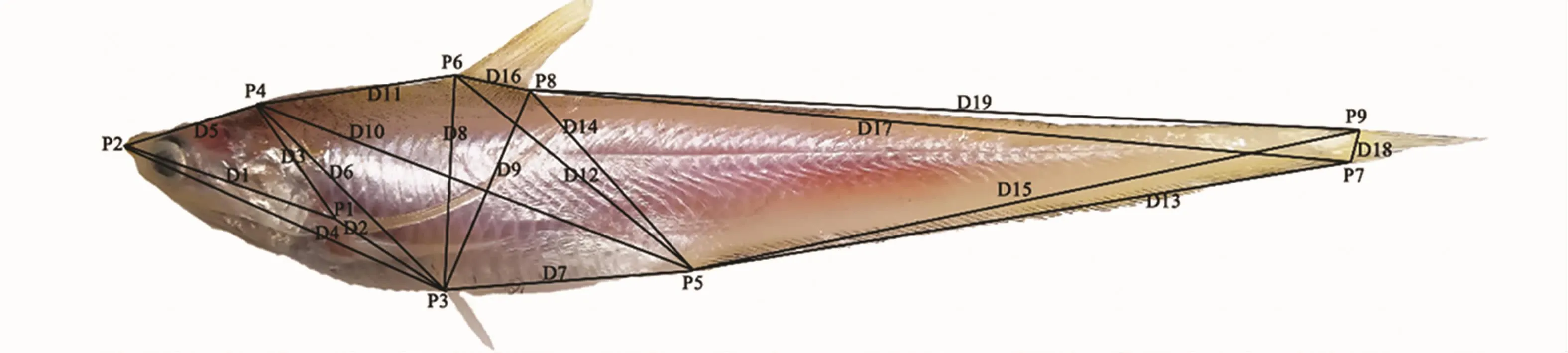
图3 凤鲚框架图Fig.3 Truss network of distance measurement of C.mystus
2 结果与分析
2.1 非参数检验
对29个比例性状进行非参数检验,Kruskal-Wallis H检验显示,除了吻长/体长(SL/BL)外,4个海域凤鲚群体间其它形态比例性状存在显著性差异(P<0.05,表2)。
2.2 主成分分析
为确定影响不同地理群体凤鲚形态差异的主成分因子,对29个形态学比例性状进行主成分分析,共获得7个主成分(PC1~PC7),累计贡献率为 86.46%,其中主成分 1贡献率为44.27%,主成分2~7的贡献率分别为16.31%、8.36%、5.51%、4.38%、4.10%和 3.53%(表3)。贡献率最大的主成分1主要显示了D7/BL、STL/BL、TL/BL、D13/BL和 D15/BL等指标,集中反映的是鱼躯干部和尾部特征;主成分2主要显示了 D18/BL、BD/BL、D8/BL、D11/BL和 D9/BL等指标,集中反映的是鱼体部和尾部特征;主成分3主要显示了 HL/BL、ED/BL、HBE/BL和 D5/BL等指标,集中反映的是鱼体头部特征。而凤鲚4个群体主成分PC1和PC2构成的散布图显示,吕四、崇明和舟山3个群体个体间交错重叠区域较多,而温州群体与其它3个群体个体重叠区域较少,分化最为显著(图4)。

表2 凤鲚4个群体的形态比例性状描述性统计值(平均值±标准差)Tab.2 Descriptive statistics of morphological proportional traits for four populations of C.mystus(Mean±SD)
2.3 判别分析
利用逐步判别和交互验证方法对29个形态比例性状进行判别分析,建立由贡献率较大的8个特征值构成的判别方程式,这8个变量分别为:FL/BL(X1)、HD/BL(X2)、TL/BL(X3)、CPH/BL(X4)、D9/BL(X5)、D13/BL(X6)、D16/BL(X7)、D19/BL(X8)
吕四群体(Ls):
YLs=38 542.907X1+3 257.822X2-29 310.853X3-9 782.854X4+1 731.135X5+25 849.984X6+5 899.037X7+15 330.884X8-24 730.902

图4 凤鲚4个群体第1、2主成分散布情况Fig.4 Distribution of PC1 and PC2 for four populations of C.mystus
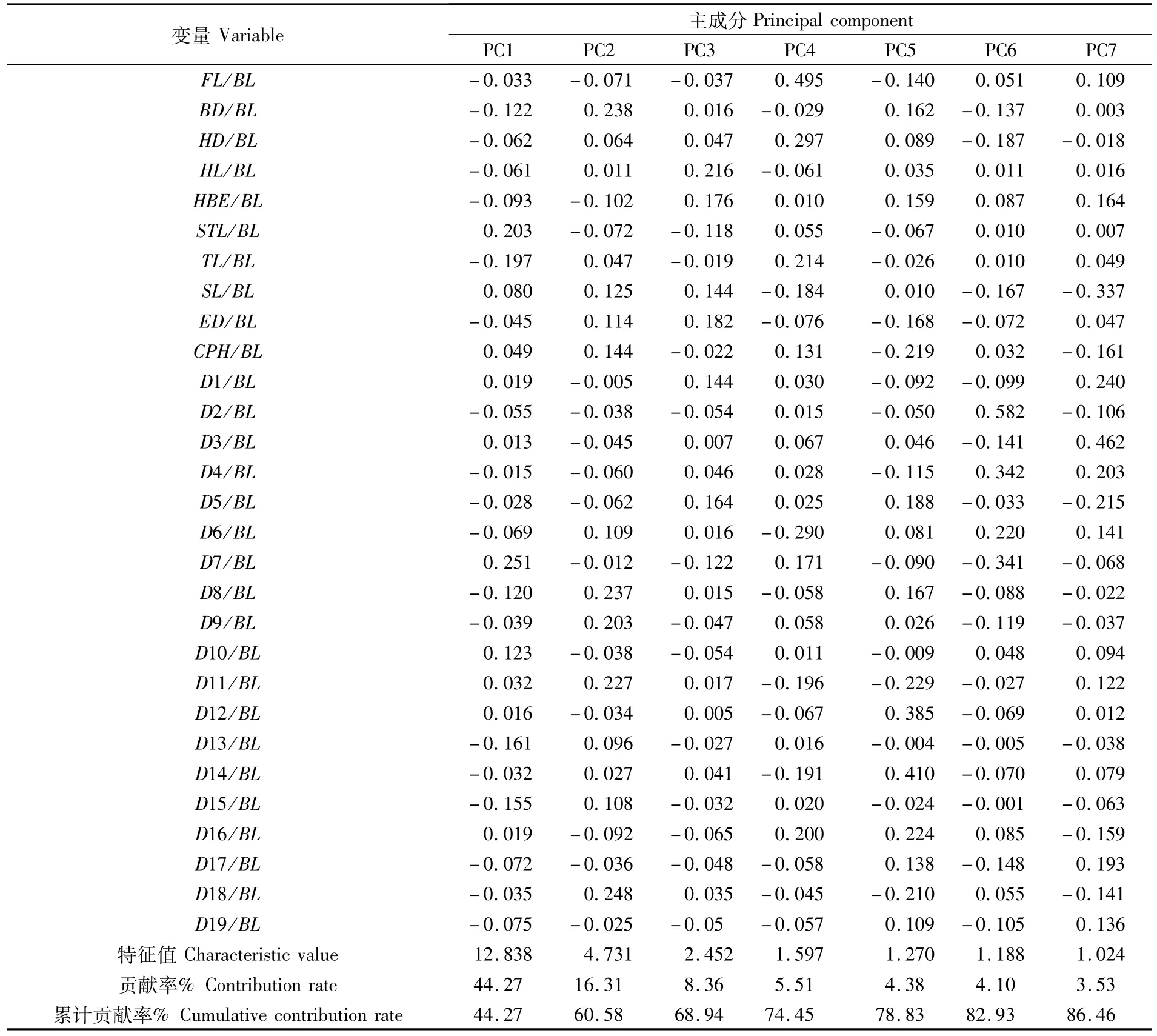
表3 凤鲚4个群体前7个主成分特征值和贡献率Tab.3 Initial eigenvalues and contribution ratio of the first seven principal components for four populations of C.mystus
崇明群体(Cm):
YCm=38 641.147X1+3 375.915X2-29 267.519X3-10 184.118X4+1 690.028X5+26 028.774X6+5 774.139X7+15 153.911X8-24 720.982
舟山群体(Zs):
YZs=38 711.580X1+3 190.348X2-29 321.998X3-9 971.162X4+1 776.692X5+26 133.413X6+5 709.9 36X7+15 158.544X8-24 814.329
温州群体(Wz):
YWz=38 464.330X1+3 029.771X2-29 241.006X3-9 319.513X4+1 865.921X5+25 928.780X6+5 548.9 22X7+14 980.518X8-24 356.985
逐步判别分析结果显示:综合判别准确率为76.3%,4个群体判别成功率依次为温州群体(90.0%)、吕 四 群 体 (87.5%)、崇 明 群 体(67.5%)和舟山群体(60.0%)。交互验证分析结果显示:综合判别准确率68.1%,吕四、崇明和舟山群体判别率分别低于逐步判别分析结果,而温州群体判别率达到95.0%(表4)。
由4个群体的典型判别分析散点图(图5)可以看出,4个凤鲚地理群体个体基本是互相分离的,吕四群体集中分布在第1象限,温州群体集中分布在第2、3象限,而崇明和舟山群体都分布在第4象限,两者组质心距离最近,部分个体发生混淆。由此可见,崇明和舟山群体形态最为接近,吕四次之,温州群体与其它3个群体之间差异最大。

图5 凤鲚4个群体形态测量特征的判别分析Fig.5 Discriminant function analysis for morphological trait measurements of four population of C.mystus
2.4 聚类分析
选择29个比例性状变量的平均值,采用组间连接方法,得到群体间欧式距离树状图(图6)。从凤鲚4个地理群体间的欧式距离可以看出(表5):崇明群体(Cm)和舟山群体(Zs)形态较为接近,两者之间的欧式距离最短,仅为0.016;从聚类分析图(图6)可见:凤鲚4个群体被分为两大类群,其中崇明和舟山群体距离最近,最先聚合为一组,后与吕四群体汇为一大组,而温州群体单独聚成一组。由此可见,崇明和舟山群体形态最为相似,而与吕四和温州群体的形态差异逐渐增大。

图6 由欧式距离产生的凤鲚4个群体的聚类分析图Fig.6 Cluster analysis of four populations of C.mystus in the Euclidean distance
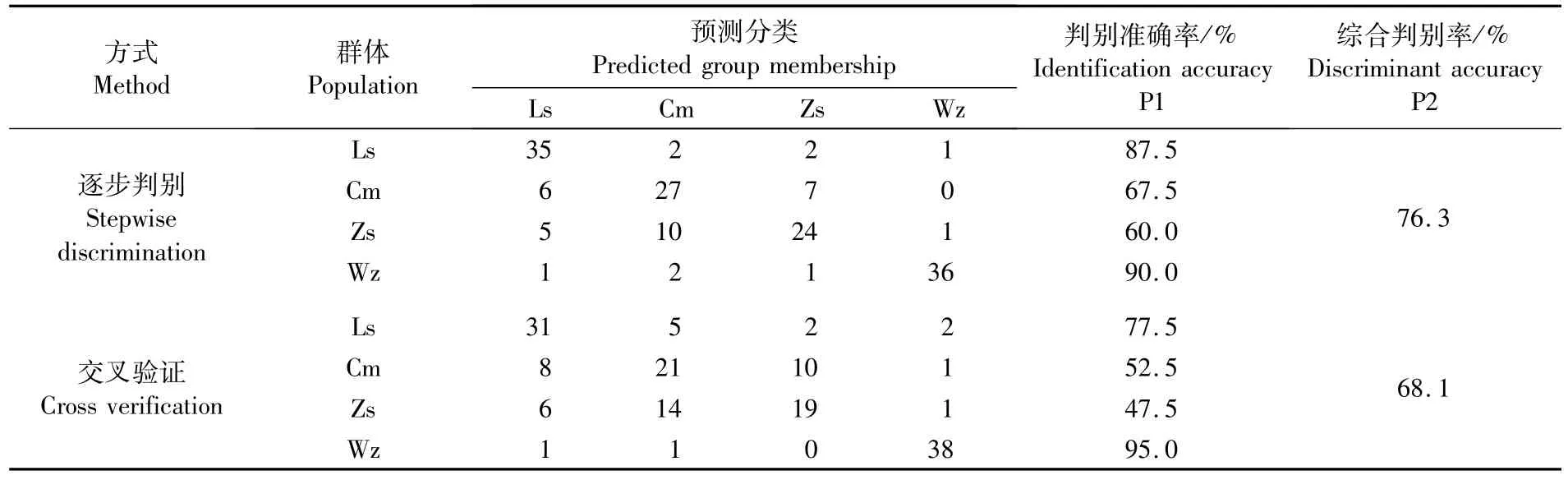
表4 基于8个贡献显著的比率变量判别函数对4个群体凤鲚判别分析结果Tab.4 Results of discriminant analysis of four populations of C.mystus based on four discriminant functions with eight significant ratio variables

表5 凤鲚4个地理群体间的欧式距离Tab.5 Euclidean distance among four geographical populations of C.mystus
3 讨论
长期地理隔离以及对不同环境的适应性使得同种鱼类不同地理群体间由于缺乏基因交流而产生一定的生殖隔离,从而在其形态、生理及遗传上形成一定的差异[13,19-20]。早期研究表明,我国东南沿海凤鲚群体存在显著的地理分化[21],可分为长江型、闽江型和珠江型3个生态类群[22],群体间的分化可能达到了亚种水平[23-24]。刘文斌等[25]基于生化和形态学研究,将长江口凤鲚和九龙江凤鲚划分为2个亚种。由此可见,地理区域相距较远的凤鲚群体之间存在较大的形态差异,然而凤鲚作为一种短距离洄游性鱼类,大部分凤鲚群体的生殖、索饵、育肥等过程都在既定的区域内进行,大体不会超出其生存环境的范围,推测地理位置相距较近的凤鲚群体之间也可能存在形态差异。
本研究中,Krushal-Wallis H检验和主成分分析直观有效地判断出长江口及邻近海域凤鲚4个地理群体间的总体关系,除了吻长/体长外,其它形态比例存在显著性差异,主成分分析将多个形态比例性状转化为几个相互独立或互不相关的几个因子[26],根据主成分贡献率大小,得到影响群体差异的主要形态参数,共提取了7个主成分,然而前3个主成分的累积贡献率较低,仅为68.94%,未达到累积贡献率大于或等于85%的要求,说明凤鲚群体难以用几个相互独立的因子概括不同群体的形态差异[27]。其中主成分1贡献率最大,为44.27%,主要反映了 D7/体长、躯干长/体长、尾长/体长、D13/体长和 D15/体长等鱼体躯干部、尾部横向和斜向的形态差异。棘头梅童鱼(Collichthyslucidus)[27]、刀鲚[13]和红鳍东方鲀[14]等鱼类的研究也表明躯干部和尾部特征是影响不同地理种群差异性的主要因素,这与鱼类游泳能力是相符合的,鱼类游泳主要通过躯干和尾部肌肉强有力的活动而快速前进,其产生的动力远远大于鳍条划水产生的动力。从主成分分析的散点图来看,温州群体分布较为特殊,与其它3个群体重叠率较小,而吕四、崇明和舟山3个群体之间重合度较高,分布较为集中,形态差异较小,由此推断这些差异性大小与4个群体所处地理位置有关,崇明群体处于长江口海域,北靠吕四群体所处的黄海南部渔场,南靠舟山群体所处的东海舟山渔场,这3个群体生存的自然环境、气候以及水体都极为相似,从而导致了3个群体的形态相似性,而温州群体处于东海入海河流瓯江北部,与其它3个群体相隔较远,地理的阻碍以及栖息地环境的不同导致群体间缺乏基因交流,从而产生较大的群体差异,可单独划分为一个群体。
聚类分析和判别分析结果较为一致,都有效地对4个凤鲚群体进行了判别及分类,崇明和舟山群体首先聚为一组,再与吕四聚为一大组,温州群体独自为一组,且崇明和舟山两个群体组质心和欧式距离最为接近,清楚地显示了群体间的亲缘关系及关联性,即崇明和舟山群体亲缘关系较为接近,而与吕四和温州群体亲缘关系逐渐增大。通过具有筛选变量能力的逐步判别分析组合最佳因子建立了稳定的判别函数[28],对4个不同地理群体凤鲚进行了判别,只有吕四和温州群体判别准确率较高,错判率在15%以内,说明可以有效地进行判别,根据2个群体的地理位置,推测其分别属于黄海入河口洄游群体和瓯江洄游群体。而崇明和舟山群体判别较低,2个群体个体混合程度较高,与主成分分析结果一致,崇明和舟山群体所处海域存在长江和钱塘江2个相近的入海口,相近的水文体系使得2个群体间交流频繁,群体内个体出现交叉,推测2个群体都属于长江洄游型或钱塘江洄游型群体;值得注意的是吕四群体判别率较高,这与和其地理位置相近的崇明和舟山群体之间存在较为显著的差异,与主成分分析结果不一致,与其地理位置分布也不相符合,分析可能是由于台风、洋流等的输送作用下,黄海北部黄海入河口洄游群体具浮游性的卵及幼体扩散至黄海南部,与长江型洄游群体混淆,从而造成了这种差异。
综合以上研究结果,采用4种多元统计学方法从不同角度说明了4个不同地理凤鲚群体之间既存在差异又存在密不可分的关联性,证实了长江口凤鲚群体组成以崇明群体及舟山群体为主,可能存在少数来自长江口以北的黄海水域迁移的吕四凤鲚群体,因此今后对长江口凤鲚资源保护管理单元的划分应集中在长江以南和舟山以北的东海水域为主。本研究在前人基础上更精确和全面分析了长江口及邻近海域凤鲚群体差异性,为长江口凤鲚种质资源保护区域划分及管理提供了有效的理论依据,然而不同地理群体凤鲚形态差异也有可能是由遗传因子、环境因素及自然选择相互作用造成的[28-29],与群体间的遗传、生理等有关,所以今后要对凤鲚种质资源做更全面深入的评估,并需从凤鲚群体耳石元素指纹、微卫星标记、生理生化及分子生物学等方面做进一步比较研究。
致谢:中国水产科学研究院东海水产研究所河口渔业实验室纪严、徐超、徐静静和吴晓峰同学在取材与实验方面给予了帮助,谨致谢忱。
