间作条件下超积累和非超积累植物对重金属镉的积累研究
2019-07-01霍文敏范洪黎
霍文敏,邹 茸,王 丽,范洪黎*
[1.中国农业科学院农业资源与农业区划研究所/农业农村部植物营养与肥料重点开放实验室,北京 100081;2.中国地质大学(北京)土地科学技术学院,北京 100083]
据报道,当前我国约有1/5的耕地存在重金属污染,其中Cd污染耕地面积为1.33万hm2。有关研究表明,8种土壤重金属元素中Cd污染概率达25.2%,远远高于其他重金属元素[1]。White等研究表明,Cd在植物组织中的浓度达到5~10 mg/kg就会对植物体造成毒害[2]。由于Cd化合物的溶解度高于其他金属化合物,因此Cd在水-土壤-植物系统中具有很强的迁移能力[3],且能够通过食物链在人体中积累[4],美国毒物管理会(ATSDR)将Cd列为人体健康的有毒物质的第6位[5]。植物提取修复技术是利用超积累植物来修复污染土壤或水体的一种成本低、有效的土壤Cd修复技术[6-7],但由于超积累植物大多生长缓慢、生物量小、大范围推广难度大,因此急需筛选富集能力强、生物量大、适应性好的重金属Cd修复植物[8]。
目前,国内外发现的Cd超积累植物有17种,且多数为草本植物[9]。魏树和等通过室外盆栽模拟试验及重金属污染区试验,首次发现并证实杂草龙葵(Solanum nigrum L.)是一种Cd超积累植物[10]。龙葵具有抗逆性强、生长迅速以及在环境适宜条件下能够急剧提高等特点,同时,当Cd浓度为25 mg/kg时,龙葵茎、叶中Cd含量分别为103、124 mg/kg,地上部Cd富集系数为2.68[11]。黑麦草(Lolium perenne L.),多年生植物,具有根系发达、适应性强、生长快、地上部分生物量大、可多次收割的特点[12-13],因而在用作重金属修复植物资源方面具有非常大的潜力。杨卓等通过温室盆栽试验表明,种植黑麦草3个月后,地上部、根部Cd含量分别为5.57、114 mg/kg[14],监测黑麦草体内重金属含量的变化发现,地上部分对Cd具有超强的富集能力[15]。
目前,国内外Cd污染土壤的植物修复中,对于超积累植物的研究取得了一定的进展[16],但是对于间作条件下超积累及非超积累植物不同器官的生长及对Cd吸收转运特性的研究还比较少[17]。本文以超积累植物龙葵和非超积累植物苋菜、黑麦草为研究对象,采用温室盆栽试验,研究玉米间作条件下对3种植物的生长,对Cd的吸收转运机制以及对Cd污染土壤的修复效率,为今后筛选Cd污染土壤中最适宜间作的植物以及提高土壤的修复效率提供依据。
1 材料与方法
1.1 供试土壤
采自河南省北部某市,该地区属温带大陆性季风气候,年平均气温14℃,全年日照时数约2 400 h,年平均降水量656 mm。从污染点的农田进行采土作为供试土壤,该供试土壤质地为棕壤砂土,采多点进行混合,土壤风干、磨碎后分别过2、0.25 mm筛,土壤理化性质及重金属含量的测定结果见表1。
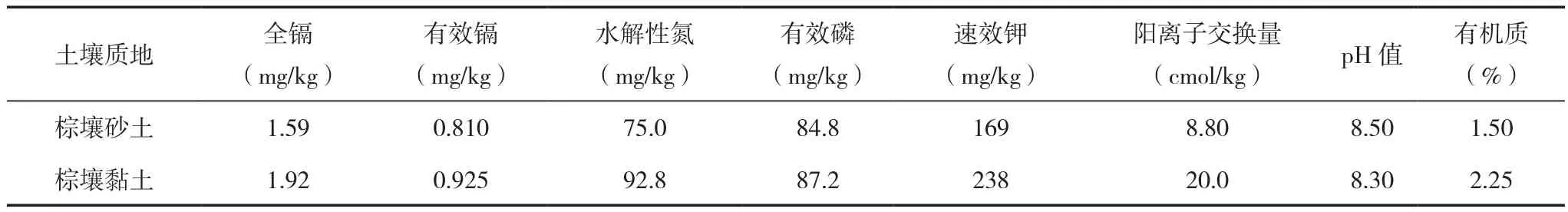
表1 供试土壤基本理化性质
1.2 供试作物
供试玉米由河南省农业科学研究院提供,品种为郑单958号。玉米种子经10% H2O2溶液消毒20 min,然后用去离子水冲洗几遍,在培养皿中用去离子水浸种催芽24 h,待种子发芽后播种至土壤中。供试超积累植物选用野生龙葵,采用种子育苗,龙葵种子先经温水浸泡12 h,然后用湿毛巾包起来置于25℃以上催芽,待幼苗长出两片真叶后选择大小相近,长势一致的幼苗移栽,每盆定植幼苗4株。供试非超积累植物选用苋菜、黑麦草。
1.3 试验设计
盆栽试验于2017年6月在中国农业科学院温室中进行,采用白色PVC圆盆(直径35 cm,高30 cm)[18-19],将供试Cd污染土装盆,每盆装10 kg土,将催芽后的玉米种子、龙葵幼苗、苋菜种子、黑麦草种子同时移入盆中,分别设置以下7种处理,重复3次。7种处理分别为玉米单作(CK)、苋菜单作、黑麦草单作、龙葵单作、玉米-苋菜间作、玉米-黑麦草间作、玉米-龙葵间作。每盆移栽玉米1株,间作植物龙葵、黑麦草、苋菜均为4株。其中,盆中央移栽1株玉米,距四周以10 cm,4株间作植物等间距分布。植株统一在20~30℃温室条件下培养,生长期间每1~2 d进行浇水,保持在田间持水量的70%左右。玉米、龙葵、苋菜、黑麦草4种植物生长期一致,为2017年6月15日至9月30日。
1.4 样品采集与指标测定
收获时,植物成熟期采集土壤样品和植株样品。每盆用小型不锈钢土钻随机采取5点,组成1个混合土样。土壤样品自然风干磨碎后分别过2、0.25 mm筛备用;植株样品分根、茎、叶和籽粒4部分取样,清水洗净后再用去离子水冲洗,在105℃烘箱中杀青30 min,80℃烘干,同时测生物量,并将根、茎、叶和籽实分别磨碎,过0.25 mm筛备用。
植物中Cd前处理:取过0.25 mm筛的植株样品 0.25 g,采用 HNO3-HC1O4(4∶1)消解,定容至25 mL;土壤Cd全量前处理:取过0.25 mm筛的土壤样品0.20 g,采用王水-H2O2消解,定容至50 mL待测;土壤有效态Cd前处理:称取过2 mm筛孔的风干盆栽土壤5.00 g放入100 mL锥形瓶中,加入DTPA提取剂25.0 mL,在往复振荡器上振荡提取2 h。振荡频率为(180±20)r/min,振荡温度为(25±2)℃。弃去最初2~3 mL滤液,滤液待测[14]。DTPA提取剂的制备:称取1.97 g DTPA(二乙三胺五乙酸)溶于14.9 g(13.3 mL)TEA(三乙醇胺)和少量水中,再将1.11 g CaCl2溶于水中,一并转入1 000 mL容量瓶中,加水至约950 mL,用6 mol/L HCl溶液调节pH值至7.30(每升提取液约需加6 mol/L HCl溶液8.50 mL),最后用水定容,贮存于塑料瓶中[20]。
Cd含量均采用电感耦合等离子体质谱法测定,电感耦合等离子体质谱仪ICP-MS为美国Agilent 7700x。测定过程中质控样品为国家标准物质GBW10020(GSB-11),Cd标准值为(0.17±0.02)mg/kg,测定值为0.18 mg/kg,质控样品测定结果在标准值范围内。
1.5 富集系数与转运系数
富集系数(BCF)=玉米地上部Cd含量/土壤中Cd元素含量
转运系数(TF)=玉米地上部Cd含量/地下部Cd元素含量
1.6 统计分析
所有数据的平均值、标准差采用Excel 2007软件,数据的方差分析、相关性分析及聚类分析采用SPSS 19.0软件。试验数据用SPSS 19.0统计分析,多重比较用Duncan新复级差法。字母不同表示差异显著(5%水平),字母相同者表示不显著。
2 结果与分析
2.1 间作条件下不同超积累植物与非超积累植物的生物量
本试验中选取的几种富集植物的种类不同,各器官的生物量也存在差异,具体如表2所示。在整个试验过程中,3种间作植物龙葵、黑麦草、苋菜生长良好,未出现任何中毒现象,因此,这3种植物都具有较强的耐Cd特性。地上部生物量最大的为龙葵,其次是苋菜,黑麦草的生物量最小。其中,土壤Cd浓度为1.59 mg/kg时,龙葵地上部生物量分别是苋菜、黑麦草的2.41、10.6倍;土壤Cd浓度为1.92 mg/kg时,龙葵地上部生物量分别是苋菜、黑麦草的2.42、9.06倍。方差分析显示,3种富集植物地上部生物量的差异达到显著水平(P<0.05)。
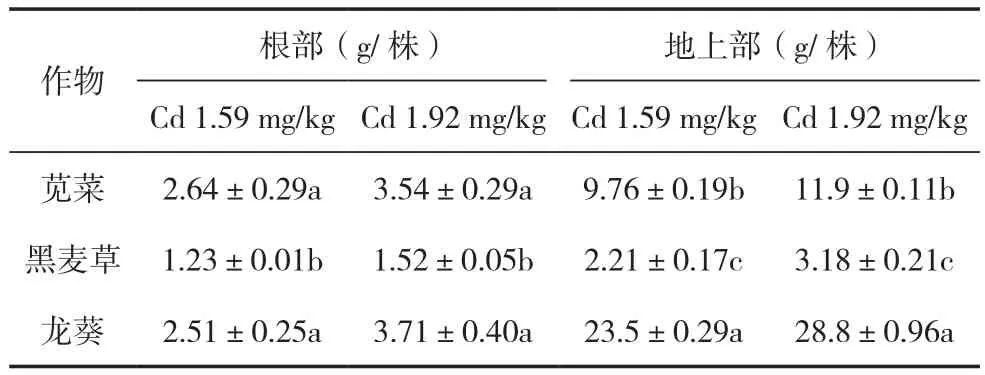
表2 超积累植物、非超积累植物与玉米间作下的生物量
2.2 间作条件下不同超积累植物与非超积累植物对Cd含量的影响
由图1可知,间作条件下超积累植物龙葵各器官中Cd含量表现为:叶>茎>籽粒>根,即地上部大于根部,表明超积累植物向地上部转运能力强;而苋菜中Cd含量表现为:根>叶>茎,黑麦草中Cd含量表现为:根部>地上部,即非超积累植物Cd含量为根部大于地上部。土壤Cd污染程度越高,间作植物苋菜、黑麦草、龙葵对重金属Cd的吸收积累作用越强。其中,黑麦草根部Cd含量最高,其次是苋菜,龙葵根部Cd含量最低;龙葵地上部Cd含量最高,其次是苋菜,黑麦草地上部Cd含量最低。Cd浓度为1.59 mg/kg时,龙葵茎、叶中Cd含量分别是苋菜的18.8、10.4倍;Cd浓度为1.92 mg/kg时,龙葵茎、叶Cd含量分别是苋菜的12.0、9.97倍,表明超积累植物龙葵地上部对Cd的吸收积累远远大于非超积累植物苋菜、黑麦草。
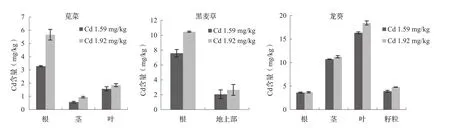
图1 间作下超积累植物与非超积累植物的Cd含量
2.3 间作条件下超积累植物和非超积累植物对Cd累积量的影响
不同超积累植物、非超积累植物间作下对Cd积累的影响如图2所示,间作条件下超积累植物、非超积累植物根部Cd累积量的大小顺序为:黑麦草>龙葵>苋菜,而地上部Cd累积量的大小顺序为:龙葵>苋菜>黑麦草。Cd浓度为1.59、1.92 mg/kg时,龙葵地上部Cd累积量分别为273、367μg/株,远大于超积累植物的临界标准[21]。其中,Cd浓度为1.59 mg/kg时,龙葵地上部Cd累积量分别为苋菜、黑麦草的28.0、59.9倍;Cd浓度为1.92 mg/kg时,龙葵地上部Cd累积量分别为苋菜、黑麦草的30.8、43.5倍,且超积累植物龙葵与非超积累植物苋菜、黑麦草的地上部Cd累积量差异显著(P<0.05)。因此,超积累植物龙葵对Cd的累积量远大于非超积累植物苋菜、黑麦草。
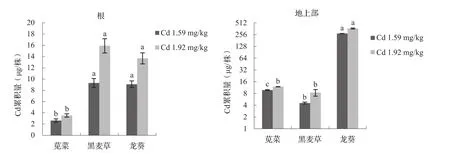
图2 间作条件下超积累植物与非超积累植物Cd累积量的差异
2.4 间作条件下不同超积累植物和非超积累植物对土壤Cd全量、有效态Cd含量的影响
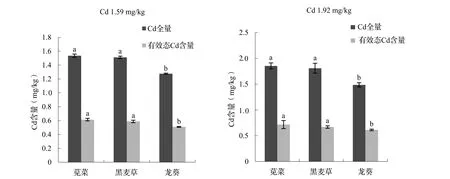
图3 间作条件下超积累植物与非超积累植物对土壤Cd全量、有效态Cd含量的影响
玉米间作条件下不同超积累、非超积累植物对土壤中Cd全量、有效态Cd含量的影响不同,其中,间作苋菜的土壤中Cd全量及有效态Cd含量最高,其次是黑麦草,间作龙葵的土壤中Cd全量及有效态Cd含量最低,表明龙葵间作模式对Cd污染的土壤修复作用最强。土壤中Cd含量为1.59 mg/kg时,龙葵间作后的土壤Cd全量显著性下降,苋菜、黑麦草、龙葵与玉米间作下土壤Cd全量分别下降了3.46%、4.97%、19.8%;土壤中Cd含量为1.92 mg/kg时,苋菜、黑麦草、龙葵与玉米间作下土壤Cd全量分别下降了3.43%、5.79%、22.6%,其中龙葵间作模式下显著性下降。土壤中Cd含量为1.59 mg/kg时,苋菜、黑麦草、龙葵间作后的土壤有效态Cd含量分别下降了24.1%、27.4%、37.1%;土壤中Cd含量为1.92 mg/kg时,苋菜、黑麦草、龙葵间作后的土壤有效态Cd含量分别下降了22.9%、28.1%、34.1%。
2.5 间作条件下不同超积累植物和非超积累植物的富集系数、转运系数
由表3可以看出,土壤中Cd含量为1.59 mg/kg时,间作模式下超积累植物龙葵的富集系数是苋菜、黑麦草的28.2、59.3倍,转运系数是苋菜、黑麦草的8.08、55.9倍,差异达到显著水平。土壤中Cd含量为1.92 mg/kg时,龙葵的富集系数分别是苋菜、黑麦草的29.4、41.4倍,转运系数分别是苋菜、黑麦草的7.98、53.6倍。间作条件下超积累植物龙葵的富集系数、转运系数远大于1,而非超积累植物苋菜、黑麦草的富集系数均小于1,表明非超积累植物苋菜、黑麦草对Cd的积累能力较低。综上可知,超积累植物龙葵对土壤中Cd的吸收与转运能力远大于非超积累植物苋菜、黑麦草。
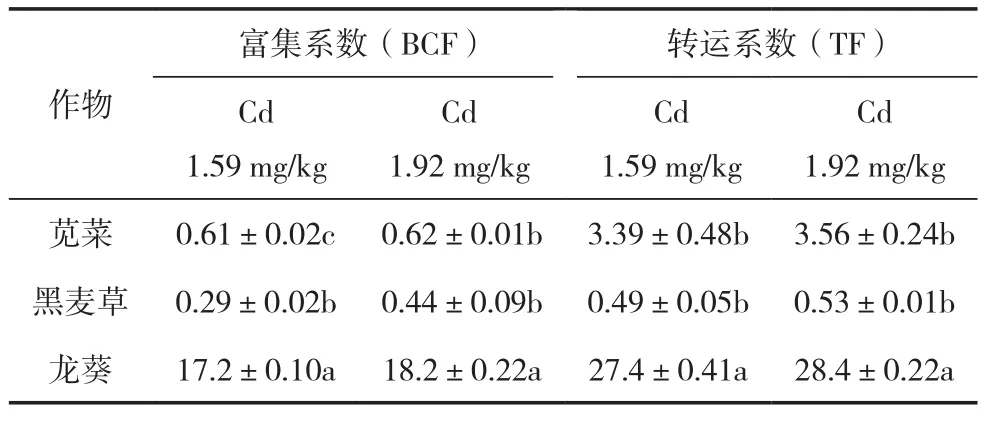
表3 间作条件下超积累植物与非超积累植物对Cd的富集系数、转运系数的差异
3 讨论
在同一污染Cd浓度下,不同间作模式下植物地上部Cd累积量差异显著,地上部Cd积累量的大小顺序为:龙葵>苋菜>黑麦草,即Cd胁迫下,超积累植物龙葵对Cd的累积量远远大于非超积累植物苋菜、黑麦草。由表3可知,不同Cd污染浓度下,苋菜、黑麦草对Cd的富集系数均小于1,而龙葵对Cd的富集系数均大于16,这与熊国焕等研究结果相似[22]。富集系数能够反映元素在土壤-植物这一系统中的吸收能力,表明间作模式下超积累植物龙葵对Cd的积累能力远远大于非超积累植物苋菜、黑麦草。有关研究表明,植物对重金属Cd的吸收过程中,吸收速率(Vmax)起到关键的作用[23]。Cd超积累生态型N.caerulescens Granges对Cd的最大吸收速率约为Prayon生态型的5倍[24],A.halleri与S.alfredii两种超积累植物也表现出相似的结果[25-26]。
研究表明,超积累植物和非超积累植物地上部对Cd的吸收积累存在差异,龙葵地上部Cd累积量远高于非超积累植物苋菜、黑麦草。Cd污染浓度分别为1.59、1.92 mg/kg时,龙葵地上部Cd累积量分别为273、367μg/株。该结果与黑亮等一致,黑亮等通过研究玉米与超积累植物东南景天套种,发现东南景天提取Zn的效率显著提高[27]。间套种条件下,由于玉米一侧根际土壤中pH值降低,水溶性有机物(DOC)提高,使得土壤中水溶性Cd浓度增加,从而促进更多的水溶性Cd向间作作物一侧移动,由于超积累植物龙葵能够吸收更多的重金属Cd并富集在体内[28],因此超积累植物Cd累积量显著高于非超积累植物。这一现象其原因与超积累植物特殊的解毒机制有关[29],超积累植物对重金属的解毒过程主要有2个途径:一是合成一些小分子化合物与重金属形成络合物,降低其毒性;二是将重金属区隔于活性较低的部位,如液泡、细胞壁等[30-31]。而对于非超积累植物,其根系吸收Cd2+后,一部分在根系细胞壁和液泡中积累,另一部分通过根细胞的跨膜运输进入木质部,再通过茎运输至叶片中[32]。
研究表明,不同Cd污染浓度处理下,龙葵各器官Cd含量分布表现为根>茎>籽粒>叶,即地上部>根部,这一研究结果与魏树和等一致[10]。苋菜、黑麦草Cd含量分布为根部>地上部,即非超积累植物Cd主要积累在地下部。由表3可知,不同Cd污染浓度下,黑麦草对Cd的转运系数均小于1,而龙葵对Cd的转运系数均大于27。因此,超积累植物龙葵对Cd的转运能力显著高于非超积累植物苋菜、黑麦草。对于这一结果,是由于超积累植物龙葵的根系向地上部转运重金属Cd的能力强,而超积累植物根系向地上部高效的重金属转运过程主要由两方面的过程控制:一是超积累植物根细胞对重金属的截留作用小,二是超积累植物具有高效的木质部装载过程[33]。超积累植物能够尽可能减少重金属在根系细胞液泡的积累,并且能够将液泡里面存储的重金属最快速度地转运到细胞质进而使其能够往地上部转运[34]。Cd非超积累植物相对于超积累植物来说,根系向地上部的转运效率低很多,可能与其根细胞液泡对Cd的截留作用强有关[35]。
4 结论
玉米间作模式下,从超积累植物和非超积累植物的生物量来看,地上部分生物量的表现为:龙葵>苋菜>黑麦草,黑麦草的生物量最小。其中,土壤Cd浓度为1.59 mg/kg时,龙葵地上部生物量分别是苋菜、黑麦草的2.41、10.6倍;土壤Cd浓度为1.92 mg/kg时,龙葵地上部生物量分别是苋菜、黑麦草的2.42、9.06倍。玉米间作模式下,超积累植物龙葵各器官中Cd含量表现为:叶>茎>籽粒>根,即地上部大于根部;而苋菜中Cd含量表现为:根>茎>叶,黑麦草中Cd含量表现为:根部>地上部,即非超积累植物Cd含量为根部大于地上部。土壤中Cd含量为1.59 mg/kg时,龙葵地上部Cd累积量分别为苋菜、黑麦草的28.0、59.9倍;龙葵的富集系数分别是苋菜、黑麦草的28.2、59.3倍,转运系数分别是苋菜、黑麦草的8.08、55.9倍,差异达到显著水平;苋菜、黑麦草、龙葵间作后的土壤Cd全量显著性下降,分别下降了3.46%、4.97%、19.8%。土壤中Cd含量为1.92 mg/kg时,龙葵地上部Cd累积量分别为苋菜、黑麦草的30.8、43.5倍,且超积累植物龙葵与非超积累植物苋菜、黑麦草的地上部Cd累积量差异显著;龙葵的富集系数分别是苋菜、黑麦草的29.4、41.4倍,转运系数分别是苋菜、黑麦草的7.98、53.6倍;苋菜、黑麦草、龙葵间作后的土壤Cd全量分别下降了3.43%、5.79%、22.6%,其中龙葵间作模式土壤Cd全量呈显著性下降。综上可知,超积累植物龙葵对土壤中Cd的吸收与转运能力远远大于非超积累植物苋菜、黑麦草,是玉米间作模式中最理想的植物修复材料。
