绵羊KISS1基因组织表达及其多态性与产羔数关联分析
2019-06-18安雪姣赵生国潘章源文禹粱田志龙储明星
安雪姣,赵生国,潘章源,文禹粱,田志龙,蔡 原*,储明星*
(1.中国农业科学院北京畜牧兽医研究所,农业部动物遗传育种与繁殖重点实验室,北京 100193;2.甘肃农业大学动物科学技术学院,甘肃兰州 730070;3.临沂大学农林科学学院,山东临沂 276005)
KISS1 基因最初是Lee 等从人黑色素瘤细胞中分离得到,具有抑制肿瘤转移的作用,被定位于人的1q32-41[1-2]。KISS1 基因的翻译产物Kisspeptin 是由145 个氨基酸组成的亲水性蛋白,具有分泌神经肽的特点,是G 蛋白偶联受体54(GPR54)的配体,并组成KISS1/GPR54 系统[3],该系统在哺乳动物下丘脑-垂体-性腺轴(HPG)繁殖调节中起中枢调控作用[4]。研究表明,给非繁殖季节的绵羊体内注射Kisspeptin 能显著提高促黄体素(LH)和促性腺激素释放激素(GnRH)水平,从而导致绵羊排卵[5]。说明KISS1 基因能直接调节LH 和GnRH 的合成与分泌,从而促进动物的季节性繁殖活动[5-6]。
我国大多数绵羊品种属于季节性繁殖,仅有少数品种为常年发情。小尾寒羊是我国特有的优良地方品种,一年四季均可发情繁殖。草地型藏羊属于季节性繁殖绵羊,一般在秋冬发情繁殖。本研究分别采集小尾寒羊和草地型藏羊10 种繁殖相关组织样品,检测KISS1 基因在这2 个绵羊品种不同组织中的表达差异,并对重测序筛选出来的KISS1 基因g.1317523C>T 和 g.1311578G>T位点在6 个绵羊品种中的多态性进行分析,初步分析KISS1 基因在绵羊季节性繁殖中的作用,以及其多态性与绵羊产羔数之间的关系,为进一步开展绵羊季节性繁殖的研究和分子标记辅助育种提供参考依据。
1 材料与方法
1.1 实验样品 基因表达实验:随机选择生长状况良好、饲养时间相同(2~3 周岁)的西藏当雄县的草地型藏羊和山东省郓城县的小尾寒羊经产空怀母羊各3 只。屠宰后,迅速采集大脑、小脑、下丘脑、垂体、卵巢、子宫、输卵管、肾脏、甲状腺、肾上腺共10 种组织新鲜样品,装入2 mL 冻存管,液氮中保存,转移到-80℃冰箱保存备用。
多态性分析实验:21 只苏尼特羊血液样本采自内蒙古巴彦淖尔市乌拉特中旗民羊牧业有限责任公司,22 只滩羊血液样本采自宁夏盐池县宁夏朔牧盐池滩羊繁育有限公司,161 只草地藏羊血液样本采自西藏当雄县,101 只湖羊血液样本采自江苏省徐州申宁羊业有限公司,52 只策勒黑羊血液样本采自新疆和田市策勒县,380 只小尾寒羊血液样品采自山东省郓城县诚联小尾寒羊种羊场。采用颈静脉采血,柠檬酸葡萄糖抗凝-20℃保存。同时记录小尾寒羊产羔季节、胎次与产羔数。
1.2 组织总RNA 的提取和检测 使用Trizol(Invitrogen,美国)和动物组织总RNA 提取试剂盒(天根生化有限责任公司,北京)提取藏羊和小尾寒羊各组织总RNA。用NanoDrop 2000 检测所提取RNA 的浓度及纯度(A260/A280在1.8~2.1),用1%的琼脂糖凝胶电泳检测RNA 完整性,置于-80℃保存备用。
1.3 引物设计 根据GenBank 绵羊KISS1 基因mRNA 序列(登录号:NM_001306104.1)使用Primer Premier 5.0软件跨外显子设计引物,以β-actin(GenBank 登录号:NM_001009784)作为内参基因。引物由北京天一辉远生物科技有限公司合成。引物具体信息见表1。

表1 绵羊KISS1 基因的引物信息
1.4 cDNA 合成和qPCR 参考周梅等[7]、李晓雨等[8]的cDNA 合成方法和qPCR 体系和程序,绘制目的基因和持家基因的标准曲线,检测引物的扩增效率,并确保其在95%~105%,进行qPCR 反应以检测目的基因的相对表达量。
1.5 基因分型 采用Sequenom MassARRAY®SNP 技术[9]对KISS1 基因g.1317523C>T 和 g.1311578G>T 位点进行基因型检测。分型样品为DNA,每个样品需要量为20 μL,DNA 浓度为40~80 ng/μL。
1.6 统计分析 实时荧光定量数据分析 :采用2-△△CT法[10-11]计算目的基因相对表达量,用SPSS 19.0 统计软件对数据进行单因素方差分析,结果用平均值±标准误表示,以P<0.05 表示差异显著。
基因分型数据分析参考本课题组田志龙等[12]的分析方法用Excel 2016 软件统计绵羊KISS1 基因2 个SNPs 的基因型频率、等位基因频率、多态信息含量(PIC)、杂合度(He)和有效等位基因数(Ne),然后进行Hardy-Weinberg 平衡检验,用SPSS 19.0 软件对小尾寒羊基因型与产羔表型数据关联进行分析,结果用平均值±标准误表示。
2 结果与分析
2.1 总RNA 提取与cDNA 合成 提取小尾寒羊和草原型藏羊各组织的总RNA,浓度均在100 mg/μL 以上,A260/A280均在1.8~2.1。用1%琼脂糖凝胶电泳检测,28 S 和18 S条带清晰可见,28 S 亮度高于18 S,且无拖带现象,说明提取的总RNA 无明显降解(图1)。经反转录合成的cDNA 以持家基因β-actin 进行RT-PCR 扩增后,产物干净单一(图2),表明提取的RNA 和合成的cDNA 可用于后面的qPCR 试验。

图1 不同品种绵羊10 种组织RNA 电泳检测

图2 不同品种绵羊10 种组织中β-actin 持家基因RT-PCR 扩增产物
2.2 KISS1 基因在小尾寒羊和草地型藏羊中的表达 如图3 所示,KISS1 基因在2 个绵羊品种的10 种组织均有一定表达,在肾脏、肾上腺和输卵管中表达量很低。小尾寒羊下丘脑、大脑、垂体和甲状腺组织中KISS1基因的表达量高于草地型藏羊(P<0.05)。
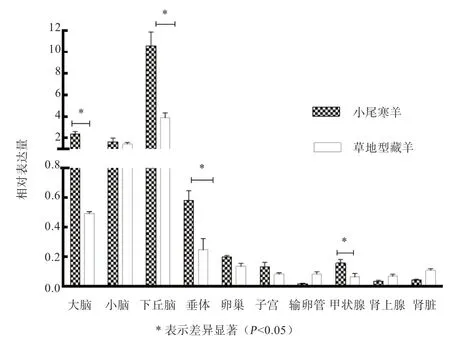
图3 KISS1 基因在小尾寒羊和草地型藏羊各组织间的表达
2.3 KISS1 基因多态性分析 如图4 所示,KISS1 基因g.1317523C>T 位点在常年发情和季节性发情绵羊品种中均存在3 种基因型,分别是CC、CT 和TT;g.1311578G>T 位点在常年发情和季节性发情绵羊品种中也存在3 种基因型,分别是GG、GT 和TT。
由表2 可知,绵羊KISS1 基因g.1317523C>T 位点基因型频率和等位基因频率在常年发情和季节性发情绵羊品种间差异均达到极显著水平(P<0.01),且无论在常年发情还是季节性发情品种中C 均是优势等位基因;g.1311578G>T 位点基因型频率和等位基因频率在季节性发情、常年发情绵羊品种间差异不显著,且在季节性发情和常年发情绵羊品种中G 均是优势等位基因。
从表3 可知,绵羊KISS1 基因g.1317523C>T 位点在小尾寒羊、滩羊、苏尼特羊、策勒黑羊、湖羊和草原型藏羊6 个群体中均表现为中度多态(0.25<PIC<0.5);卡方适合性检验结果表明,该位点在小尾寒羊、滩羊、苏尼特羊和湖羊4 个群体中均处于哈代温伯格平衡状态(P>0.05),而在草地型藏羊和策勒黑羊中处于不平衡状态。g.1311578G>T 位点在小尾寒羊、滩羊、策勒黑羊、湖羊和草地型藏羊5 个绵羊群体中表现为低度多态(PIC<0.25),在苏尼特羊中表现为中度多态(0.25<PIC<0.5);卡方适合性检验结果表明,该位点在苏尼特羊和湖羊品种中均处于哈代温伯格平衡状态(P>0.05),而在滩羊、小尾寒羊、草地型藏羊和策勒黑羊中均处于不平衡状态。

图4 KISS1 基因g.1317523C>T(A)与g.1311578G>T(B)的分型结果
2.4 KISS1 基因2 个SNPs 位点与小尾寒羊产羔数的关系 由表4 可知,KISS1 基因2 个SNPs 位点多态性与小尾寒羊不同胎次产羔数之间并无显著相关(P>0.05),但从整体产羔水平来说,g.1311578G>T 位点TT 型产羔数大于GT 和GG 型。
3 讨 论
3.1 KISS1 基因表达分析 KISS1 基因在啮齿动物和哺乳动物的季节性繁殖调控中发挥重要作用[13],在人、猪、山羊等多个组织广泛表达[14-16]。小尾寒羊和阿勒泰羊中的研究发现,KISS1 基因在绵羊下丘脑中的表达变化有明显的周期性,其中在发情期的表达量极显著高于其他阶段(发情前期、发情后期和间情期)[17-18]。KISS1 基因mRNA 在雌性小鼠下丘脑中的性发育早期表达很高,并在性成熟期到达最高峰,表明KISS1 基因在雌鼠的性发育早期开始分泌,从而激活GnRH 的合成与分泌[19]。在常年发情山羊(济宁青山羊和金堂黑山羊)与季节性发情山羊(辽宁绒山羊和藏山羊)中的研究发现,KISS1 基因在下丘脑中的表达均显著高于其他组织,且在常年发情山羊下丘脑的表达量显著高于季节性发情山羊[16,20]。本研究结果与以上研究结果相似,在小尾寒羊(常年发情)下丘脑、大脑、垂体和甲状腺组织的表达量均显著高于草地型藏羊(季节性发情)。说明下丘脑KISS1 基因的差异表达可能是引起小尾寒羊和草地型藏羊季节性繁殖差异的重要原因之一。推测该基因在绵羊季节性繁殖调控中起重要作用。

表2 KISS1 基因2 个SNPs 位点在季节性发情、常年发情绵羊品种中的基因型频率和等位基因频率

表3 KISS1 基因2 个SNPs 位点在不同绵羊品种中的群体遗传学分析
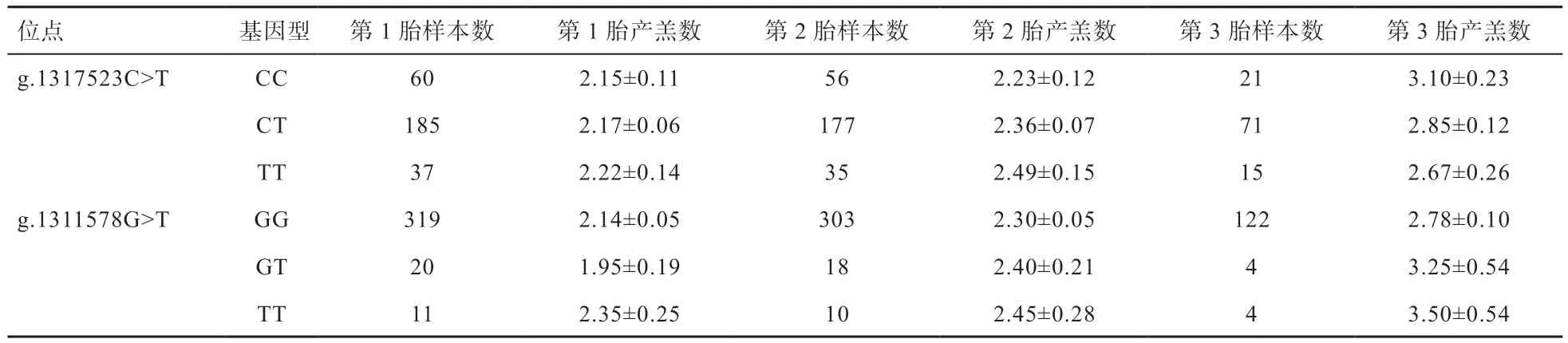
表4 KISS1 基因2 个SNPs 位点各基因型小尾寒羊产羔数最小二乘均值及标准误
3.2 KISS1 基因多态性与动物繁殖之间的关系 目前,有关KISS1 基因的多态性研究主要集中在人的中枢性性早熟[21]以及动物产羔性状[22]等方面,而KISS1 基因的多态性与动物季节性繁殖关系的相关研究未见报道。本研究发现,KISS1 基因的SNPs 位点(g.1317523C>T)在不同繁殖特性(季节性发情和常年发情)绵羊群体的基因型频率和等位基因频率均达到极显著水平,且常年发情的绵羊群体等位基因T/C 比值(0.79)高于季节性发情的绵羊群体(0.35)。表明该位点T 等位基因与绵羊常年发情性状具有相关性,推测该位点突变对绵羊的季节性繁殖有一定的影响,但其具体的影响机制有待进一步证实。
金慧慧[23]研究发现,将猪KISS1 基因转进小鼠体内,会在小鼠下丘脑内检测到KISS1 基因的表达,并且转基因小鼠每窝产仔数均高于野生型小鼠,推测KISS1基因的表达可能对小鼠产仔数有影响。An 等[24]在萨能奶山羊、波尔山羊和关中奶山羊3 个群体KISS1 基因的内含子1 上均检测出了2 个SNPs 位点(G2124T>A和G2270C>T),且都与山羊的产羔数显著相关。Martia 等[25]在印度山羊KISS1 基因内含子1 上发现了3 个与山羊产羔数相关的SNPs 位点(G296C、G2510A和C2540T)。陈祥等[26]在黔北麻羊KISS1 基因内含子1 中发现的2 个SNPs 位点(G277C、T486A)与其产羔数显著相关。本研究在小尾寒羊KISS1 基因上检测到了2 个SNPs 位点,其多态性与小尾寒羊不同胎次产羔数均无显著相关,但g.1311578G>T 位点TT 型产羔数大于GT 和GG 型,推测该位点突变在一定程度上增加了小尾寒羊的产羔能力。
4 结 论
本研究发现,KISS1 基因在小尾寒羊(常年发情)下丘脑、垂体、大脑和甲状腺的表达量显著高于草地型藏羊(季节性发情)。绵羊KISS1 基因g.1317523C>T位点与季节性发情显著相关,可作为常年发情绵羊选育的辅助标记,而g.1311578G>T 位点对绵羊产羔性状选育有潜在调控作用。
