眼部情绪信息对疼痛共情影响的ERP 研究 *
2019-06-15魏艳秋解鸿宇胡亚奇
宋 娟 魏艳秋 杜 梦 解鸿宇 连 涛 胡亚奇 马 欧
(1 教育部人文社会科学重点研究基地天津师范大学心理与行为研究院,天津 300074) (2 天津师范大学心理学部,天津 300074) (3 国民心理健康评估与促进协同创新中心,天津 300074) (4 上海师范大学教育学院,上海 200234) (5 陕西师范大学心理学院,西安 710062) (6 华东师范大学心理与认知科学学院 200062)
1 引言
共情是个体理解和分享他人情绪情感和思维的过程,在人类社会互动中占有重要地位(Decety& Jackson, 2004)。Gladstein(1983)提出了认知共情和情绪共情的概念。而Davis(1983)在其人际反应指针问卷(Interpersonal Reactivity Index,IRI)中将共情分为观点采择(perspective-taking)、幻想(fantasy)、共情关注(empathy concern)、个人苦恼(personal distress)。和Davis 观点有相似之处的研究总结发现,共情可被分解为多个成分:经验分享,指的是间接地感受他人的经验;心智化能力,涉及到积极推测他人的想法和目的(Cameron, Spring, & Todd, 2017);怜悯之心,包含减轻他人遭遇的一种动机(Decety & Cowell,2014)。共情在社会生活中有着非常重要的作用(岳童, 黄希庭, 2016),被认为是亲社会行为的一种合适的解释(Ma, Wang, & Han, 2011),有利于照顾后代、与人合作和社会交往等活动。
有研究总结发现,这20 年共情研究的主题从早期的人格特质、态度、情绪,逐渐转向了社会认知(颜志强, 苏彦捷, 2017)。而且随着研究技术手段的发展,有关共情的认知神经科学方面的研究应运成为现在乃至未来的热点。并且,疼痛共情是共情在社会认知神经科学领域的焦点(Singer& Lamm, 2009)。有研究认为认知共情涉及内侧前额叶皮层(medial prefrontal cortex, mPFC)、颞上沟(superior temporal sulcus,STS)、颞极(temporal poles)、腹内侧皮层(ventromedial cortex,VM cortex);情感共情包含杏仁核(amygdala)、脑岛(insula)、额下回(inferior frontal gyrus,Decety & Ickes, 2009)。其中,前中扣带回皮层和脑岛是疼痛共情中的“共享神经回路”(Danziger, Faillenot, & Peyron, 2009)。同时,镜像神经元在共情中占有重要地位,是社会沟通的“神经桥梁”(叶浩生, 2016)。
在人类日常生活中,作为整体的面孔可提供重要的社会信息,反映环境中的潜在变化(Yamada& Decety, 2009; Wang et al., 2012)。通过观察面部表情,人们可即时接受他人情绪信息,进而理解他人的内在感受。可见,面部表情与共情有着密不可分的关系:当区分面部表情时,共情特质和LPC(600-800 ms)的平均波幅显著正相关。高共情特质的被试能将注意力更多地放到面孔上(Choi& Watanuki, 2014)。同样,高共情特质的被试对面孔的总注视时间更长。且相比于中性面孔,被试对疼痛面孔有更快的早期注意定向(颜志强, 王福兴, 苏彦捷, 2016)。而以往关于疼痛共情的ERP 实证研究大多使用图片或视频来描述身体的某一部分经历疼痛或呈现疼痛的面部表情来探索当被试观察到他人疼痛时的生理指标和行为变化。疼痛图片和非疼痛图片之间的P3 差异波在消极情绪启动之下的差异波幅大于平静和积极情绪启动之下的差异波幅,且平静和积极情绪启动之下的差异波幅无显著差异(Meng et al., 2012);相比高兴表情,恐惧表情诱发了更大的ERP 波幅(Luo, Feng, He, Wang, & Luo, 2010)。相比于中性表情,痛苦表情诱发波幅更大的N1(Han, Luo, &Han, 2015)。李昂(2013)发现,愤怒和恐惧表情启动之下,疼痛图片所诱发的N1、N2 与非疼痛图片所诱发的N1、N2 无显著差异。
研究发现,面孔加工中存在眼睛加工优势。谷莉和白学军(2014)采用眼动技术发现成人对眼睛有明显的注意偏好。其次,面孔感知中存在眼睛的特异性加工。有研究发现,识别伤心情绪时,眼睛线索作用较大(王柳生, 钱萼, 张庆, 潘发达, 2011);在对恐惧和平静表情的识别中,眉目区和口唇区的作用同等重要;在对悲伤表情的识别中,被试首次注视就集中在眉目区,而且相比于口唇区,眉目区捕获了更多的注意(Eisenbarth& Alpers, 2011)。乔建中(1989)亦有如此结论。在传达惧类、哀类情感中,“眼睛是面部最富于表情、最能传递情感的部位”,也即“眼睛是心灵的窗户”这一观念是正确的。这些研究结论为本实验中所选取的悲伤和恐惧的眼部信息在相应面部情绪的代表性上面提供了证据。如此,仅透过面孔的眉目区,即可推断他人当下的心理状态(Lee & Anderson, 2017),进而调整自己的行为方式,且截至目前,单独使用眼部刺激来研究面部表情加工还处于初级阶段(李帅霞, 李萍, 王伟, 何蔚祺, 罗文波, 2017)。
此外,还有实证研究发现,在识别面部表情时,眼睛区域传达的威胁程度与全脸相同,而嘴巴区域则与全脸有着较大的差别(Fox & Damjanvic,2006)。另外,在不同社会价值朝向(social value orientation,SVO)背景中,相比于无眼神接触,亲社会被试(prosocials)在有阈上的眼神接触的情况下会表现出更多的合作行为;在阈下的眼神接触中,亲社会被试同样会做此反应。但此时亲自我被试却表现相反(Luo, Zhang, Tao, & Geng,2016)。Baron-Cohen 发明了眼神解读的测试(“Reading the Mind in the Eyes” Test,Baron-Cohen,Wheelwright, Hill, Raste, & Plumb, 2001),认为该测试反应了在无背景或是信息有限的情况下,个体解读人们复杂情绪状态的能力,该测试为共情的一个测量方法(Montgomery et al., 2016)。
综上所述,首先,眼部信息是传达情绪和面孔加工中的重要区域,目前尚无关于传递不同情绪信息的眼部特征信息(如眼神、眉毛等)对他人疼痛共情影响的研究。其次,目前的ERP 研究评估了在时间动态特征下,消极、平静、积极情绪图片启动(Meng et al., 2012)或者自我和他人启动(Ibáñez et al., 2011)之后呈现疼痛图片,以此来观察对他人疼痛共情的皮层反应。而以往研究少有细化消极情绪来进行研究,即比较同为负性情绪的悲伤和恐惧情绪对他人疼痛共情的脑电反应。恐惧意味着危险,一种强烈的应激反应,可诱发逃避反应;而悲伤意味着柔弱无助感,一定程度上可诱发人们的亲社会行为。在这种可诱发人类迥异社会行为的情境下,个体是如何感知他人疼痛的呢?这仍然知之甚少,有待进一步探究。与共情相关的理论解释中,威胁价值假说认为,对于观察者而言,他人的疼痛是一种潜在的威胁,能够激活早期的威胁觉察系统,且消极情绪能促进人类的疼痛共情(Yamada & Decety,2009);情绪分享理论认为,个体在知觉他人的动作表情等外部信息时,会自动发生模仿行为,且此时大脑中与此情绪相应的部位会被激活,结果表现为相同形态的表征(Decety, 2002)。大量研究表明,相比平静和积极事件,人类大脑对负性事件尤其敏感,会优先处理这些事件(Wang et al.,2012; Huang & Luo, 2006)。对于疼痛的反应更可能被无意识消极情绪所影响(Yamada & Decety,2009; Ibáñez et al., 2011)。从进化的角度来看,研究消极情绪对于人类生存和社会适应有着重要的意义(Grassini, Holm, Railo, & Koivisto, 2016)。综上所述,本研究选取了悲伤、平静、恐惧三种情绪,以此探究他人悲伤、恐惧与平静眼部情绪信息对疼痛共情的影响及其电生理学方面的证据。研究假设,相比平静情绪,被试在悲伤与恐惧眼部情绪启动下会呈现出更大的N 1、N 2、P3、LPC 波幅;疼痛刺激所引起的波幅要大于非疼痛刺激所引起的波幅。
2 方法
2.1 被试
遵循自愿参与实验的原则,随机选取13 名大学生为被试。其中,男6 人,女7 人,年龄范围20~27 岁。平均年龄22.92 岁,标准差2.22。被试的视力或矫正视力正常,无脑部损伤及精神疾病史。实验开始前签署知情同意书,实验结束后给予相应报酬。
2.2 实验材料
启动刺激图片为12 张从中国情绪面孔图片库(龚栩, 黄宇霞, 王妍, 罗跃嘉, 2011)截下的成人眼部图片(6 名男性, 6 名女性)。图片用Photoshop处理,图片内容包含眼睛、眉毛部分。图片的尺寸(260 像素*115 像素)、大小容量均保持一致,并且图片均为黑白画质,灰度一致。其中,有悲伤情绪眼部图片4 张,平静情绪眼部图片4 张,恐惧情绪眼部图片4 张。16 名在校大学生(13 女,3 男,平均年龄24.56 岁,标准差1.21)对图片所表达情绪的一致性程度进行了九级评价(1=完全不能代表, 9=非常能够代表; 代表程度从1 到9 递增)。结果显示,悲伤情绪眼部图片平均值为6.69,标准差1.11;平静情绪眼部图片平均值6.59,标准差0.87;恐惧情绪眼部的图片平均值为6.23,标准差1.33。对三种图片的代表度进行单因素方差分析,结果发现三者之间差异不显著,F(2, 9)=0.18,p=0.836。说明这三种材料在代表强度上无显著差异。目标刺激图片采用手部疼痛场景图片(Wang et al., 2016)。29 名在校大学学生对88 张图片进行了再评定,采用4 级评分(0 代表完全不疼,3 代表非常疼痛)。疼痛图片疼痛等级强度(平均值2.14, 标准差0.24)和非疼痛图片疼痛等级强度(平均值0.30, 标准差0.18)差异显著(t=40.96, p<0.001)。启动刺激图片与目标刺激图片均随机配对呈现。
2.3 实验程序
本研究采用启动范式和疼痛共情任务。启动刺激为平静、悲伤、恐惧情绪眼部图片,目标刺激为疼痛图片和非疼痛图片。研究为3(眼部情绪类型: 悲伤、平静、恐惧)×2(疼痛类型: 疼痛、非疼痛)完全被试内设计。
实验包括528 个trials。正式实验前进行练习。实验流程为(见图1):首先呈现注视点400 ms,随后呈现眼部图片200 ms,空屏500~700 ms,接着呈现疼痛与非疼痛图片1500 ms。要求被试对随后出现的场景图片进行疼痛判断任务。“疼痛”按“1”键,“非疼痛”按“4”键。随后呈现空屏500 ms,作为试次间隔。
2.4 仪器及记录
本研究采用Neuroscan 公司生产的64 导新型两用E R P 系统。电极点分布为国际通用的10-20 系统。采集数据时,参考电极置于左侧乳突处,离线转换成双侧乳突的平均作为参考。接地位于FPZ 与FZ 中点处。实验在一个隔音、避光、电屏蔽的脑电室进行。记录水平眼电的电极被置于眼睛左右侧面约1.5 cm 处,记录垂直眼电的电极被置于左眼上下约1 cm 处。滤波带通为0.05-100 Hz(离线滤波为: 低通30 Hz),采样率为1000 Hz。每个电极点的头皮电阻低于5 kΩ。实验材料的呈现和行为数据的采集采用stim 软件完成。
2.5 数据处理及分析
行为数据处理及分析:使用SPSS22.0 对各条件对应的反应时及正确率进行3(眼部情绪类型:悲伤、平静、恐惧)×2(疼痛类型: 疼痛、非疼痛)的两因素重复测量方差分析。
ERP 数据处理及分析:使用scan4.5 软件对脑电数据进行离线式处理,处理步骤包括滤波、转换参考电极、去除眼电干扰、剔除伪迹等。各成分时间窗口:N1 成分(峰值波幅)选取100-200 ms,N2 成分(峰值波幅)选取200-350 ms,P3(峰值波幅)成分选取350-450 ms,LPC 成分(平均波幅)选择600-800 ms。分析时程为目标刺激呈现之前200 ms 至呈现后1000 ms(宋娟等, 2016)。以目标刺激呈现前的200 ms 作为基线水平进行校正。使用SPSS22.0 对N1、N2、P3、LPC 成分的波幅值分别进行3(眼部情绪类型: 悲伤、平静、恐惧)×2(疼痛类型: 疼痛、非疼痛)×15(电极点:F3、Fz、F4、FC3、FCz、FC4、C3、Cz、C4、CP3、CPz、CP4、P3、Pz、P4)的三因素重复测量方差分析,并采用Greenhouse-Geisser 对自由度和p 值进行校正。
3 结果
3.1 行为实验结果
各条件正确率描述统计结果见表1。对其进行重复测量方差分析,结果显示均不显著(ps≥0.100)。
各条件反应时描述统计结果见表2。对行为数据的反应时进行重复测量方差分析,结果显示,眼部情绪类型主效应显著,F(2, 24)=4.68,p=0.019,partial η=0.28。两两比较发现,悲伤眼部情绪启动下反应时(757 ms)显著小于平静眼部情绪启动下反应时(770 ms)(p=0.012)。疼痛类型主效应不显著,F(1, 12)=0.00,p=0.971。眼部情绪类型和疼痛类型交互作用不显著,F(2, 24)=2.69,p=0.119。

表1 不同实验条件下正确率的平均数(标准差)(%)
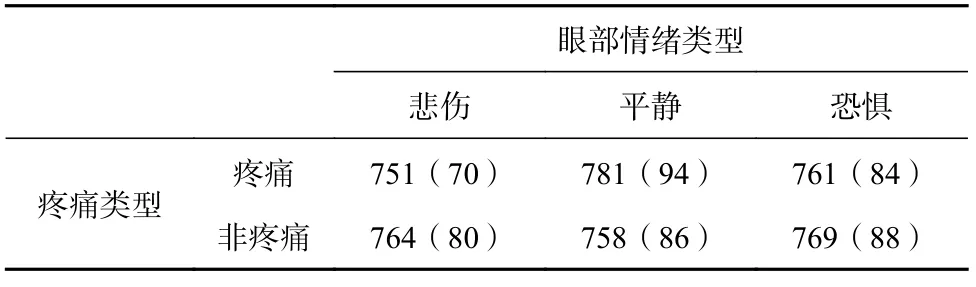
表2 不同实验条件下反应时的平均数(标准差)(ms)
3.2 电生理学实验结果
3.2.1 N1
眼部情绪类型主效应不显著,F(2, 24)=0.61,p=0.551。眼部情绪类型和疼痛类型交互作用不显著,F(2, 24)=0.08,p=0.928。疼痛类型主效应不显著,F(1, 12)=1.25,p=0.285。电极点主效应显著,F(14, 168)=19.65,p<0.001,partial η=0.62。眼部情绪类型和电极点的交互作用不显著,F(28, 336)=0.67,p=0.651。疼痛类型和电极点的交互作用不显著,F(1 4, 1 6 8)=1.1 5,p=0.343。眼部情绪类型、疼痛类型和电极点三者的交互作用不显著,F(28, 336)=1.69,p=0.173。
3.2.2 N2
眼部情绪类型主效应不显著,F(2, 24)=1.38,p=0.271。眼部情绪类型和疼痛类型交互作用不显著,F(2, 24)=3.05,p=0.066。疼痛类型主效应不显著,F(1, 12)=0.02,p=0.891。电极点主效应显著,F(14, 168)=19.64,p<0.001,partial η=0.62。眼部情绪类型和电极点的交互作用不显著,F(28, 336)=1.31,p=0.274。疼痛类型和电极点的交互作用不显著,F(14, 168)=1.72,p=0.176。眼部情绪类型、疼痛类型和电极点三者的交互作用不显著,F(28, 336)=2.08,p=0.069。
3.2.3 P3
眼部情绪类型主效应显著,F(2, 24)=4.22,p=0.027,partial η=0.26。恐惧眼部情绪的波幅(0.25 μV)小于平静眼部情绪的波幅(1.18 μV)(p=0.044),悲伤眼部情绪的波幅(0.47 μV)小于平静眼部情绪的波幅(1.18 μV)(p=0.020),恐惧眼部情绪的波幅小于悲伤眼部情绪的波幅(p=0.482)。眼部情绪类型和疼痛类型交互作用不显著,F(2, 24)=0.11,p=0.897。疼痛类型主效应显著,F(1, 12)=6.47,p=0.026,par tial η=0.35。疼痛的波幅(1.08 μV)>非疼痛的波幅(0.19 μV)。电极点主效应显著,F(14, 168)=11.68,p<0.001,partial η=0.49。眼部情绪类型和电极点的交互作用不显著,F(28, 336)=0.74,p=0.585。疼痛类型和电极点的交互作用显著,F(14, 168)=3.25,p=0.048,partial η=0.21。简单效应发现,在FC3、C3、CP3、CPZ、CP4、P3、PZ、P4 电极点,疼痛图片与非疼痛图片存在显著差异(见图2)。眼部情绪类型、疼痛类型和电极点三者的交互作用不显著,F(28, 336)=1.15,p=0.343。
3.2.4 LPC
眼部情绪类型主效应不显著,F(2, 24)=0.76,p=0.480。眼部情绪类型和疼痛类型交互作用不显著,F(2, 24)=0.71,p=0.504。疼痛类型主效应显著,F(1, 12)=15.75,p=0.002,partial η=0.57。疼痛图片引发的波幅(2.02 μV)大于非疼痛条件下的波幅(0.32 μV)。电极点主效应显著,F(14, 168)=7.73,p=0.005,partial η=0.39。眼部情绪类型和电极点的交互作用不显著,F(28, 336)=0.90,p=0.493。疼痛类型和电极点的交互作用显著,F(14, 168)=13.10,p<0.001,partial η=0.52。简单效应发现,在F3、FC3、FCZ、C3、CZ、CP3、CPZ、CP4、P3、PZ、P4 电极点,疼痛图片与非疼痛图片存在显著差异。眼部情绪类型、疼痛类型和电极点三者的交互作用不显著,F(28, 336)=1.25,p=0.290。
4 讨论
4.1 眼部情绪对疼痛共情任务的启动效应
本研究中的眼部情绪为阈上呈现,研究结果发现,三种眼部情绪在代表刺激早期加工和注意卷入程度的N1 成分上无显著差异。这一结果说明,不同眼部情绪的早期加工并无不同;同时,这三种眼部情绪在反应刺激新异程度的N2 成分上面无显著差异。这一结果和前人研究发现一致(Liddell, Williams, Rathjen, Shevrin, & Gordon, 2004;Kiss & Eimer, 2008)。在执行疼痛共情任务中,被试可能发现阈上呈现的眼部情绪与任务指示无关,因此通过自上而下的注意控制(张美晨, 魏萍,张钦, 2015)抑制住了眼部情绪对注意的捕获,从而一定程度上消除了不同类型的眼部信息在疼痛共情任务时对N1、N2 所产生的影响。眼部情绪信息在自动加工阶段没有体现出情绪对疼痛场景加工的作用,这可能是由于眼部信息的代表度和辨别度受到一定限制,需要经过进一步的评价和判断才能产生情绪效应以及情绪效应对疼痛场景判断的影响。从反应时上看,在疼痛相关物存在的场景中(疼痛/非疼痛判断任务),悲伤作为启动线索时,反应时较短,说明悲伤可产生情绪启动效应,易化此任务的靶刺激反应过程。在P3 成分上面,悲伤、恐惧眼部情绪诱发的波幅值显著小于平静眼部情绪诱发的波幅值,从某种程度上也可说明在与疼痛/非疼痛场景相关的任务中,悲伤可能与其最为相关,产生启动效应,易化对靶刺激的判断过程,减少对靶刺激注意资源的投入。恐惧眼部情绪诱发的波幅值小于悲伤眼部情绪诱发的波幅值,但差异不显著。首先,这一结果体现了大脑在P3 阶段对情绪信息的加工更为深入(张丹丹, 赵婷, 柳昀哲, 陈玉明, 2015),且在眼部情绪这一细微的局部加工上表现出差异。这说明,大脑可在情绪加工的晚期阶段区分更为精细的不同情绪类型(Campanella, Quinet, Bruyer,Crommelinck, & Guerit, 2002; Krolak-Salmon, Fischer,Vighetto, & Mauguière, 2001; Williams, Palmer, Liddell,Song, & Gordon, 2006)。其次,悲伤眼部情绪所诱发的波幅有大于恐惧眼部情绪所诱发的波幅的趋势。这一结果意味着该两种情绪的加工还是有可能对应着不同的心理过程。因为悲伤的眼部情绪更易激活人类大脑皮层的活动,人类对悲伤的情绪线索更易投入注意,进而能在一定程度上诱发疼痛/非疼痛相关场景中人类的亲社会行为(Conty,Geoge, & Hietanen, 2016);而恐惧情绪存在于一种急剧的危机情景(Kim & Cho, 2017),被试可能会基于自我保护的出发点而有意回避了对其的注意。未来研究可进一步完善眼部情绪图片对应的强度,专注比较不同眼部情绪之间的脑成分上的差异。再者,本研究的疼痛共情任务在一定程度上可能诱发了个体的焦虑心理。比如,在特定时间内让被试做疼痛判断,被试会想象自己正在遭受社会场景中主人公此时的境遇,这种任务设置可能会让被试产生一定的心理压力和情绪负担(Buffone et al., 2017),如此对原本平静的眼部情绪产生一定的负面印象和评价。故,此时对被试而言,中性情绪在一定程度上也会受任务性质的影响产生一些负性情绪效应而非单纯代表着的平静(Li, Li, Wang, Zhu, & Luo, 2017)。
4.2 疼痛共情任务
研究发现,在P3 和LPC 这两个成分中,疼痛刺激所诱发的波幅显著大于非疼痛刺激所诱发的波幅。这一结果和前人研究所得结果一致(Meng et al., 2012; Ibáñez et al., 2011; Choi et al., 2014; 杨东,李志爱, 余明莉, Ding, 姚树霞, 2015)。该结果可能与人类注意资源分配的特点有关(Meng et al.,2012)。人类注意资源是有限的,那么为了合理分配资源,人类存在负面信息偏好的注意偏差(朱永泽, 毛伟宾, 王蕊, 2014),具有进化学意义。相比于非疼痛场景刺激,疼痛场景刺激对于人类生存更具威胁性。所以,这一对生存有威胁的刺激自然会捕获更多的注意资源(Balconi & Lucchiai,2 0 0 5),接受更多的认知评估(M e n g e t a l.,2012)。因此,疼痛刺激所诱发的波幅显著高于非疼痛刺激所诱发的波幅。
5 结论
眼部表达的不同情绪信息可影响疼痛共情相关任务中的晚期加工过程。
