黄山市白背黄花稔迁移特征和种群分布格局
2019-06-02余煌辉常紫妍靳一凡曹玖玖尤宜卿王晓鹏
余煌辉, 常紫妍, 靳一凡, 曹玖玖, 尤宜卿, 王晓鹏
(安徽科技学院 生命与健康科学学院,安徽 凤阳 233100)
白背黄花稔(SidarhombifoliaL.),隶属于锦葵科黄花稔属,直立亚灌木。蒴果具8~10分果爿,种子顶端两短芒是随人或动物传播的重要结构基础。全草入药,清热利湿,消炎镇痛[1-3]。泛热带分布于非洲、亚洲南部、大洋洲、北美洲南部和南美洲,及中国的西南、中南和东南地区。杨青山等[4]首次报道白背黄花稔在安徽歙县分布新记录[4]。2017年7月在黄山屯溪郊区野外考察时发现白背黄花稔的新分布地,并对其深入调查。本研究通过分析植物区系、物种多样性、种间相关性和种群空间分布格局,以探明白背黄花稔的迁移、分布状况及其对本土植物的影响是否构成生物入侵,并为白背黄花稔资源保护与开发、评估生物多样性与生态安全提供科学依据。
1 研究地区与方法
1.1 研究区域概况

图1 安徽省白背黄花稔新记录地点和群落研究区域地理位置示意图
黄山市位于安徽省最南端,位于东经118°01'~118°17'和北纬30°01' ~30°18'之间。地处皖南山区,属中亚热带北缘常绿阔叶林地带,以山地为主。低山地大部分为黄壤、山地黄棕壤,土层较厚,石砾含量较高,透水、透气性能良好,肥力较高。属亚热带季风湿润气候区,四季分明,春秋短,夏冬长,热量丰富,雨水充沛。年平均气温15~16 ℃,无霜期236 d。年平均降水量2 398.5 mm。降水多集中于5~8月[5]。如图1所示,白背黄花稔主要集中生长在道路旁(包括公路、林间小路、田间小路等)。其样地位于屯溪郊区新潭新村与东关村相连的Y051乡道旁以及两侧丘陵的田园和林缘,各样地环境资料如表1。

表1 白背黄花稔群落各样地环境资料
1.2 样地设置与方法
共选取10个1 m2样地。2018年4月下旬调查样地Q1(幼苗期)、8月上旬调查样地Q2、Q3、Q4(旺盛生长期)、2019年10月1日调查样地Q5~Q10(跟踪加采样地)。依据不同的生境,选取Q1~Q3、Q5、Q9(1 m×1 m),Q4、Q6~Q8、Q10(2 m×0.5 m)进行野外群落调查。测度样方中植物物种数、个体数及生物量(干重),以考察群落物种丰富度指数、Simpson指数、Shannon-Wiener指数、均匀度(E)、重要值、偏离指数(Cx)、丛生指数(I)、Spearman秩相关系数检验和Sprenson系数(Ss)。
1.3 研究方法
1.3.1 物种多样性分析 物种丰富度指数(S):S为1个群落或生境(样方)中物种的数目;本研究采用α多样性(Simpson 指数、Shannon-Wiener指数和均匀度指数)和β多样性(Sprenson系数)相关指标分析群落多样性[6-7]:
1.3.2 重要值 重要值=(相对密度+相对频度+相对生物量)/3。

1.3.4 种群分布格局分析 用偏离指数的t检验法对种群空间分布类型进行划分;种群分布格局的聚集强度采用丛生指数(I)进行计算[8]:

1.3.5 植物区系分析方法 对群落中物种逐一采集标本、鉴定。蕨类植物根据秦仁昌分类系统[10]、种子植物采用恩格勒分类系统、结合模式标本及现有分布情况,根据吴征镒关于中国种子植物属的分布区类型的划分方法[11-12],对植物分布区类型进行统计分析。
1.3.6 数据处理 采用Spearman秩相关系数检验通过SPSS 16.0软件对被测数据进行单因素方差分析,得到处理结果。
2 结果与分析
2.1 白背黄花稔群落植物区系分析
2.1.1 群落植物基本组成 群落中共有植物73种,分属于32科、61属。其中,蕨类植物2科2属2种;无裸子植物;被子植物30科59属71种。尤以双子叶植物比例最大,占84.38%。其中,单属种植物有31科,占总科数96.86%,占总种数68.49%;而菊科10属13种、禾本科8属10种,分别占总种数的17.81%、13.70%。
2.1.2 群落维管植物区系成分 白背黄花稔群落维管植物包括11个属分布区类型(表2)。世界分布类型,16属,表明其植物区系中有一定数量的广域种[13]。其中,以草本植物为主,如蓼属(Polygonum)、车前属(Plantago)、酢浆草属(Oxalis)等。热带性成分共24属,占总属数的55.82%,以泛热带分布占主导地位,有18属。其中,黄花稔属(Sida)是组成群落灌木层和地被层的重要组成成分;另外,还有一点红属(Emilia)、通泉草属(Mazus)、大豆属(Glycine)、牛筋草属(Cynodon)等。温带性质成分共14属,占总属数的32.55%,次于热带性质成分;其中北温带分布占主导位,共11属,包括蒿属(Artemisia)、菵草属(Beckmannia)、风轮菜属(Clinopodium)、窃衣属(Torilis)和马兰属(Kalimeris)等。
因此,群落植物区系特征与安徽黄山植物区系特征一致[14-17],显示出由热带、亚热带向温带过渡,泛热带分布占优势的地带性区系特征。

表2 白背黄花稔群落维管植物植物区系的分布类型
注: *为百分比未包括世界分布的属
2. 2 白背黄花稔物种多样性
2.2.1 重要值 如图2所示,白背黄花稔在群落中处于优势或次优势地位,对生境异质性的适应力较强,其重要值在Q1(18.29%)、Q2(51.71%)、Q4(12.61%)、Q5(21.54%)、Q6(36.57%)、Q8(64.87%)、Q9(41.01%)、Q10(29.55%)中最大,为优势种,Q3、Q7中位居第二(15.89%、29.07%),为次优势种。其他次优势种还有酢浆草(OxaliscorniculataL.)、牛筋草(Eleusineindica)(L.)Gaertn.、空心莲子草(Alternantheraphiloxeroides(Mart.)Griseb.)、三叶委陵菜(PotentillafreynianaBornm)和早熟禾(PoaannuaL.)。

图2 白背黄花稔群落排名前五的物种重要值
Sr:白背黄花稔,Sidarhombifolia;Ds:马唐,Digitariasanguinalis;Oc:酢浆草,Oxaliscorniculata;Ei:牛筋草,Eleusineindica;Sm:繁缕,Stellariamedia;Pfl:辣蓼,Polygonumflaccidum;Cc:风轮菜,Clinopodiumchinense;As:蛇葡萄,Ampelopsissinica;Ea:飞蓬,Erigeronacer;Bs:菵草,Beckmanniasyzigachne;So:豨莶草,Siegesbeckiaorientalis;Kb:短叶水蜈蚣,Kyllingabrevifolia;Cb:荠菜,Capsellabursa-pastoris;Pfr:三叶委陵菜,Potentillafreyniana;Ap:空心莲子草,Alternantheraphiloxeroides;Gs:野大豆,Glycinesoja;Pu:叶下珠,Phyllanthusurinaria;Cy:莎草,Cyperus;Ma:鸡桑,Morusaustralis;Rh:蓬蘽,Rubushirsutus;Pn:牵牛,Pharbitisnil;Pk:蛇含委陵菜,Potentillakleiniana;Co:木防己,Cocculusorbiculatus;Pa:早熟禾,Poaannua;Pc:委陵菜,Potentillachinensis;Pp:毛竹,Phyllostachyspubescens;Ps:鸡矢藤,Paederiascandens;Sv:狗尾草,Setariaviridis;Rp:爵床,Rostellulariaprocumbens;Dc:升马唐,Digitariaciliaris;Cco:鸭跖草,Commelinacommunis
2.2.2 物种多样性分析 白背黄花稔群落各样地物种多样性均较低(图3)。其中,Q1Simpson指数(0.850 1)和Shannon-Wiener指数(3.419 1)最高;Q8的Simpson指数(0.420 0)和Shannon-Wiener指数(1.162 2)最低;从图中可看出Simpson指数、Shannon-Wiener指数和物种丰富度整体呈现近乎一致的变化趋势,即Q1、Q5、Q6、Q10的多样性指数较高,Q3、Q8处的多样性指数偏低。结合各样地生境分析:Q1透光率高、开放性程度高,存在一定强度的干扰,利于春夏之交杂草种子扩散、定居及幼苗生长,植株个体小数量大,因而物种多样性水平最高,一定程度上符合中度干扰假说[18];Q5地处弃用农田,土壤相对较肥沃,物种数较多,受干扰程度相对较低;Q6位于山坡公路的碎石地处,Q10位于051乡道旁的碎石地,共同特点是环境开放程度高,接纳物种的可能性大,四周植被较茂密,故多样性较高。Q3地处林缘,稀疏种少、富集种多,物种分布不均匀,高强度干扰和生境异质性导致种内、种间竞争加剧,故多样性较低;Q8地处田埂,由于其特殊的地势(坡度32.3°),土壤严重板结,且受人为干扰强度高,故多样性偏低。

图3 白背黄花稔群落物种多样性指数
2.2.3 样地间物种相似性分析 各样地间Sprenson系数(Ss)均小于0.5(见表3)表明各样地间物种相似性不高。Q1与Q2间Ss最高(0.33),缘于两样地间距离较近、土壤性质和生境较为接近所致;但春夏两季有不同物种占据时间生态位,因而Ss偏低。Q8与其余7个样地(Q2~Q7、Q9)间Ss为0,这可能与Q8地处坡度大(32.3°)、生境恶劣、贫瘠的田埂上有关。

表3 白背黄花稔植物群落各样地的Sprenson系数(Ss)
2.3 白背黄花稔与群落中物种种间相关性
通过Spearman秩相关系数检验,发现白背黄花稔与37个物种呈负相关,其中与马兰(KalimerisindicaL.)和三叶委陵菜呈显著负相关;与37个物种呈现正相关,其中与鳢肠(EcliptaprostrataL.)呈显著正相关(P>0.05)。总体呈不显著负相关的种对与不显著正相关的种对相持平;整个群落中各种对的种间相关性以不显著负相关性偏多,整体种间相关性较弱。
2.4 白背黄花稔种群分布格局
白背黄花稔在群落中主要呈集群分布(Cx>1)(如表4)。其中Q2、Q4~Q10的Cx,I和t≫1,呈显著的集群分布;Q1、Q3的Cx偏低接近于1,但10个样地皆满足t>t0.05,即Q1,Q3集群性较低,暗示了人为干扰、生境异质性与种间竞争等因素的影响。
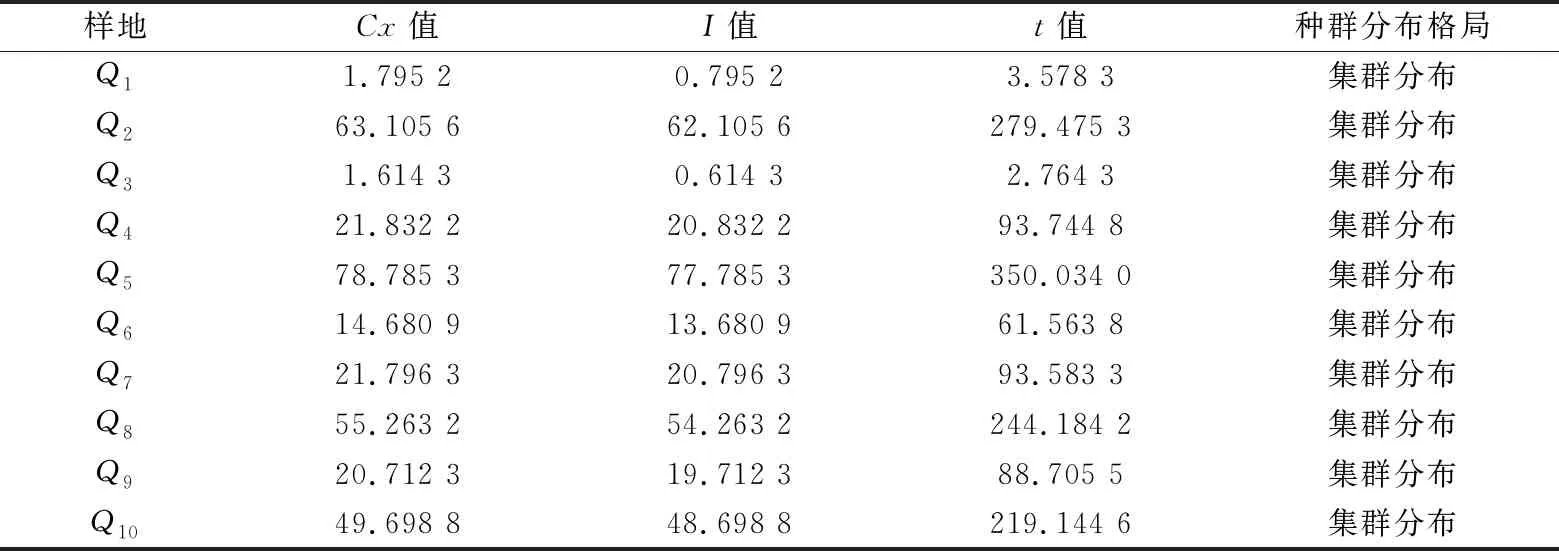
表4 白背黄花稔种群分布格局
3 结论与讨论
植物扩散的方式可分为自然扩散和人工载体扩散[19]。白背黄花稔蒴果量多,具8~10分果爿,种子小且顶端具两短芒[3],借助人工载体扩散能力较大,但自然扩散能力有限;而现有十个样地均处于Y051乡道(连接新潭新村和徽州区岩寺镇,并直达歙县新分布地[4])附近,人员和物资流动频繁,距歙县新分布地远超植物自然扩散的距离,排除自然扩散的可能[19-20];加之白背黄花稔被当地居民视为杂草,排除人为有意引入[19-20]的可能。因此,最早,可能是借助交通工具及物资交流(农作物种子、围垦建设土壤等)[21]而进行的人工载体扩散。在黄山市屯溪区分布地的地段上,扩散初期,白背黄花稔以人、动物、风力或水流等媒介[19]传播种子,使之零星分布;一旦定居,常以母株为中心向周围扩散,在适宜的生境中形成集群分布。在集群分布的前提下,由于其成体植株高大(亚灌木)、丛生,影响居民生产、生活,部分会被人为拔除,导致集群性下降。
白背黄花稔与群落中74个物种呈现或正或负的种间相关性,以不显著负相关的种对居多,与其在黄山市屯溪区的“新分布”充分契合,群落处于演替初期阶段[7,22],物种多样性处于较低水平,同时,群落中可能存在长期连续的干扰[23],且环境异质性高。各样地间物种相似性不强,反映出白背黄花稔对不同生境、重度干扰[24]和种间竞争有较强的适应能力,能与其他物种相互共存,并能保持其较高的重要值,显示其在群落中处于优势或次优势地位。在白背黄花稔定居后,与本土植物形成共存的种间关系,对生境的适应、改造[25],已基本趋于稳定。群落物种组成相对简单,区系成分缺少中国特有分布、中亚分布等分布类型,但地带性特征明显,显示出由热带、亚热带向温带过渡,泛热带分布占优势的区系性质[14-17]。群落中不乏酢浆草和辣蓼等广域种[13]分布,且马唐能在Q3中占据优势地位,均表明白背黄花稔对本土植物整体影响不显著;但其集群分布、成株个体大,对群落结构的改造以及农业生产活动存在客观的影响,也是降低群落物种多样性的原因之一。
综上所述,白背黄花稔通过人工载体扩散被偶然带入研究区域;白背黄花稔的定居对本土植物生物多样性有一定程度的影响,但远不及生物入侵的水平;此外,其生物学特性、生态学特征以及群落的脆弱程度[20]互相制约,使其仍具有转化成生物入侵的可能,因此,我们需进一步对其生态安全、风险评估进行长期监测,并适度开发和利用该植物资源[1-2],以抑制其种群大规模扩散,控制种群数量;同时,对其植物种质资源进行必要的保护,以维护该地区的生物多样性。
