糜子矮秆突变体778农艺性状调查及其对GA的敏感性分析
2019-05-21贾小平杨德智戴凌峰寇淑君张小梅侯典云朱学海
张 博,贾小平,*,杨德智,赵 渊,戴凌峰,寇淑君,张小梅,侯典云,朱学海,*
(1.河南科技大学 农学院,河南 洛阳 471000; 2.张家口市农业科学院,河北 张家口 075000)
农作物育种实践中,作物株高与抗倒伏、产量等重要农艺性状紧密相关。对植物株高进行改良,已成为创造理想株型的主要技术路线,而获得新的优异种质资源,则是株高改良中必不可少的一步。矮秆性状作为一个常见性状,曾推动了第一次“绿色革命”,使得小麦和水稻的产量得以大幅提升。优质的矮秆资源可以用来培育矮化且高产的新品种,并且可以有效解决倒伏问题[1-5]。因此,深入研究高矮秆不同农艺性状之间的差别以及探索控制株高的内在分子机制对于进一步提高作物产量至关重要。
目前对于矮秆突变体的研究较多,如王益军等[6]对一份玉米显性矮秆突变体进行了遗传分析,研究表明,玉米显性矮秆突变体D8、D9的典型突变表型为植株矮化、叶片窄而浓绿、分蘖增加、花期延迟、雌穗上着生花药、对外源赤霉素施用不敏感,矮秆基因D-10被定位在玉米第2染色体上。陈立[7]比较了玉米矮秆突变体K125d与其同源自交系K211的农艺经济性状差异,研究表明与同源自交系K211比较,K125d生育期极显著延长,穗位高极显著降低,平均株高降低53.07%,叶片重叠密集,叶片数目和叶宽极显著增加,但叶夹角极显著减小,利于理想株型建成,单株产量降低34.41%,但穗长和穗行数差异不显著,有增产潜力,其矮化原因是茎秆细胞长度缩短导致的节间长度缩短。王随宝等[8]对几个冬小麦矮秆种质材料做父本时的矮化作用进行了研究评价,鉴定筛选出陕160、西北矮个子和临5064/烟农15三种类型的优异矮秆种质,并提出了这些优异种质的利用设想。
关于矮秆基因的深入研究,也已经有相当多的报道。如小麦Rht基因,其编码的蛋白抑制了赤霉素的信号转导途径,从而导致植株矮化[9];水稻sd1基因也有很多相关报道,由于其编码的缺陷型GA20氧化酶,从而影响了赤霉素的生物合成途径,使植株高度受到影响[10];甘蓝型油菜一个半显性矮秆突变体中发现DS-1基因位于A6染色体上,编码作为赤霉素受体的DELLA蛋白[11]。对矮秆基因的研究表明,矮秆性状与赤霉素的生物合成和转导途径有关。
糜子作为一种比较抗旱的禾谷类作物,具有稳定的生产力和较好的经济效益,在旱作农业以及抗灾救灾中是其他作物不可替代的[12-17]。关于糜子的研究多是关于抗病虫害以及产量构成因素的分析,而对于其矮化突变体的诱导以及相关基因挖掘至今未见相关报道[18-23]。因此,本研究从一份糜子矮秆突变体材料778入手,对糜子苗期进行形态学观察以及7个田间农艺性状进行测定,并通过喷施赤霉素处理来进一步探索其矮化原因是否与赤霉素生物合成和转导途径相关,为进行控制突变性状的基因克隆和功能分析奠定基础。
1 材料与方法
1.1 植物材料
实验于2017年在河南科技大学农场进行,材料为糜子原始高秆材料260,以及矮秆突变材料778(由高秆260经EMS诱变),材料种子均由张家口农业科学院朱学海研究员提供。
1.2 实验药品和仪器
KH2PO4,KNO3,Ca(NO3)·4H2O,MgSO4·7H2O,H3BO3,MnCl·4H2O,ZnSO4·7H2O,CuSO4·5H2O,NaMoO4·2H2O,FeSO4·7H2O,Na-EDTA,无水乙醇,以上试剂均为国产分析纯。赤霉素为美国Sanland公司生产的分析纯试剂。光照培养箱GZX-150B由上海坤天仪器有限公司生产、微量移液器由Eppendorf公司生产。
1.3 材料培养和处理
1.3.1 田间种植及管理
糜子矮秆突变体778、原始高秆260于2017年5月18日播种于河南科技大学开元校区试验田,株距5 cm,行距40 cm,每品种种植3行,行长2 m,生长期间对田间进行标准化管理,在糜子生育期拍照记录表型形态。
1.3.2 室内植物材料和生长状态
将实验材料种子置于4 ℃冰箱48 h,之后用4个塑料小盆(10 cm×10 cm)装一半泥土,每个小盆种植150粒种子,2盆原始材料,2盆矮秆突变材料,置于恒温培养箱中,每日浇水等待萌发。等待材料生长至四叶期,进行水培移植,水培方法参照Hoagland方法进行,每3 d更换一次培养液。
1.3.3 赤霉素喷施处理
使用50、100、200 mg·L-1赤霉素溶液,分别喷施不同处理组,对照组用水进行喷施。以上处理间隔6 d喷施一次。
1.4 表型数据的测定
1.4.1 田间表型数据的测量
糜子成熟后,在试验田随机选取长势均一糜子矮秆突变体778、糜子原始高秆材料260各10株进行株高、穗长、节间长、节间数、种子长、种子宽和穗粒数这7个田间农艺性状的测定。如果有分蘖,测主茎的农艺性状。
1.4.2 赤霉素处理组数据的测量
从赤霉素处理植株开始,每隔6 d对赤霉素实验组和对照组株高、根长进行测量并记录数据。
1.5 统计分析
将记录好的数据用Excel处理,并计算各浓度下高秆材料260与矮秆突变材料778的GA敏感系数。利用SPSS17.0进行单因素的方差分析,并用邓肯法进行多重比较分析。
2 结果与分析
2.1 矮秆突变体的外观形态分析
如图1-a和b所示,在苗期,矮秆突变体778和原始材料高秆260的株高已出现明显差异。矮秆突变体778的地上基部节和第一、第二伸长节均短于原始高秆材料260。此外矮秆突变体的叶片短而宽,叶色深绿,而高秆材料的叶片细长,叶色较浅。成熟后高秆材料的穗较长,且松散,而矮秆材料的穗短,较紧实;高秆材料从穗下第一节开始,5个节间长度均大于矮秆突变材料。
2.2 矮秆突变体的农艺性状分析
由表1可以看出,矮秆突变体778的穗长为11.35 cm,高秆260的穗长为24.58 cm,差异极显著(P<0.01)。矮秆突变体778的节间数为4.7个,高秆260的节间数为5.5个,差异极显著(P<0.01)。矮秆突变体778的种子长为0.30 cm,高秆260的种子长为0.25 cm,差异显著(P<0.05)。矮秆突变体778和高秆260的种子宽差异不显著(P>0.05),高秆260的种子宽略大于矮秆突变体778(图2)。矮秆突变体778的穗粒数和株高分别为129.9粒和75.5 cm,高秆260的穗粒数和株高分别为523粒和124.6 cm,差异极显著(P<0.01)。

a,苗期矮秆突变体778(右侧)和原始材料高秆260(左侧)株高比较;b,成熟后矮秆突变体778(右侧)和原始材料高秆260(左侧)穗与节间长比较。a, Comparison of plant height of dwarf mutant 778 (the right) and the original high straw 260 (the left); b, Comparison of spikes and internode length of mature dwarf mutant 778 (the right) and original high straw 260 (the left).图1 原始高秆260和矮秆突变体778的外观形态比较Fig.1 Comparison of the appearance of the original high straw 260 and the dwarf mutant 778
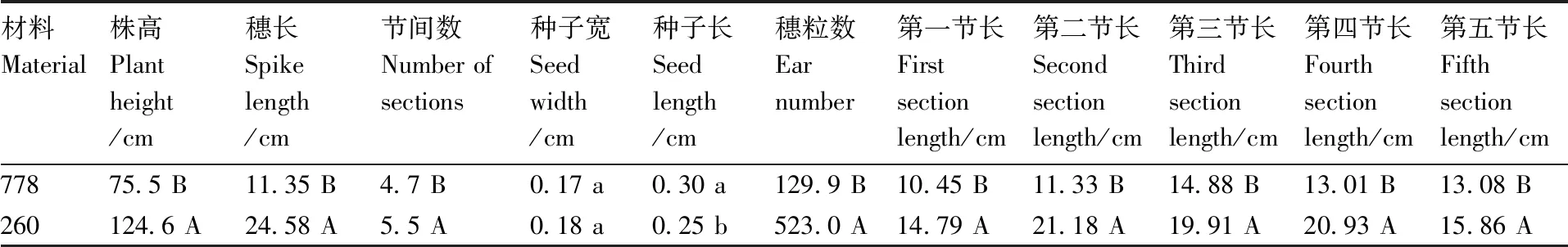
表1 两种材料部分表型数据
同列不同行数据后没有相同小写字母表示差异显著(P<0.05),没有相同大写字母表示差异极显著(P<0.01)。
The data in the same column without the same lowercase or capital letters showed the significant different at the level of 0.05 or 0.01, respectively.
矮秆突变体778的第一节间长度、第二节间长度、第三节间长度、第四节间长度、第五节间长度分别为:10.45、11.33、14.88、13.01、13.08 cm,高秆260的第一节间长度、第二节间长度、第三节间长度、第四节间长度、第五节间长度分别为:14.79、21.18、19.91、20.93、15.86 cm,差异均达到极显著水平(P<0.01)。
2.3 赤霉素敏感性分析
根据表2可以看出来,矮秆材料在3个赤霉素处理浓度下株高均大于空白对照,且随着时间的延长,处理组较对照组株高差异变大,处理组株高明显大于对照组株高;从第12天(第2次测量)开始,随着GA浓度的增加,矮秆突变体株高也呈现递增的趋势。矮100的GRI值大于矮50的GRI值(GRI=1.15、1.14、1.21)且略大于矮200的GRI值(GRI=1.08、1.25、1.43),说明矮秆突变体对于100 mg·L-1的赤霉素浓度更为敏感。根据表中数据可以看出,矮秆突变体在经过3次赤霉素喷施后,其株高可以恢复至高秆正常株高,说明导致矮秆突变的原因可能与赤霉素合成途径有关。
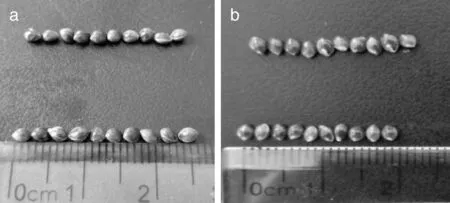
a,种子长度比较;b,种子宽度比较。a, Comparison of seed length; b, Comparison of seed width.图2 矮秆突变体778(上部)和高秆260(下部)穗粒比较Fig.2 Grain comparison of dwarf mutant 778 (the upper) and high stalk 260 (the lower)
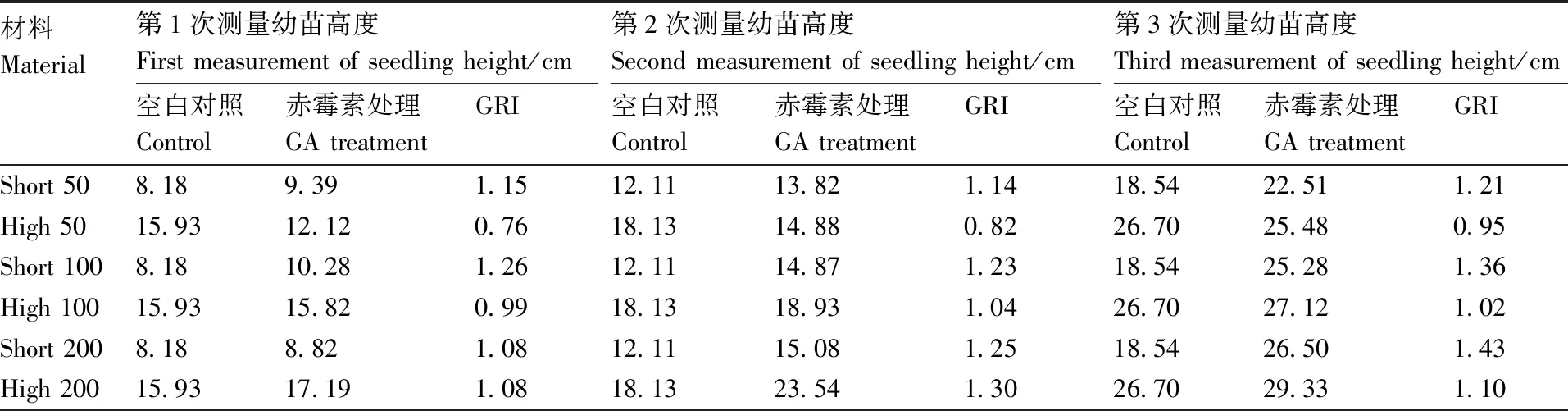
表2 高秆材料260与矮秆突变材料778的幼苗高度及GRI值
Short 50、Short 100、Short 200:矮秆突变体778分别用50、100、200 mg·L-1GA处理;High 50、High 100、High 200:高秆260分别用50、100、200 mg·L-1GA处理。第1次测量的时间为赤霉素处理后的第6天;第2次测量的时间为赤霉素处理后的第12天;第3次测量的时间为赤霉素处理后的第18天。
Short 50, Short 100, Short 200: Dwarf mutant 778 was treated with 50, 100, 200 mg·L-1GA; High 50, High 100, High 200: High straw 260 was treated with 50, 100, 200 mg·L-1GA, respectively. The first measurement time was the sixth day after gibberellin treatment, the second measurement time was the twelfth day after gibberellin treatment, and the third measurement time was the eighteenth day after gibberellin treatment.
2.4 不同质量浓度赤霉素对糜子单个性状的影响
由图3-A可以看出,第6天和第12天的矮秆材料株高在赤霉素为0、50、100、200 mg·L-1不同组之间差异不显著(P>0.05)。第6天时,赤霉素200 mg·L-1组的高秆材料的株高要显著高于赤霉素50 mg·L-1组(P<0.05),其他组之间无显著差异(P>0.05)。第12天时,赤霉素200 mg·L-1组的高秆材料的株高要极显著高于赤霉素50 mg·L-1组(P<0.05),显著高于其他组(P<0.05)。
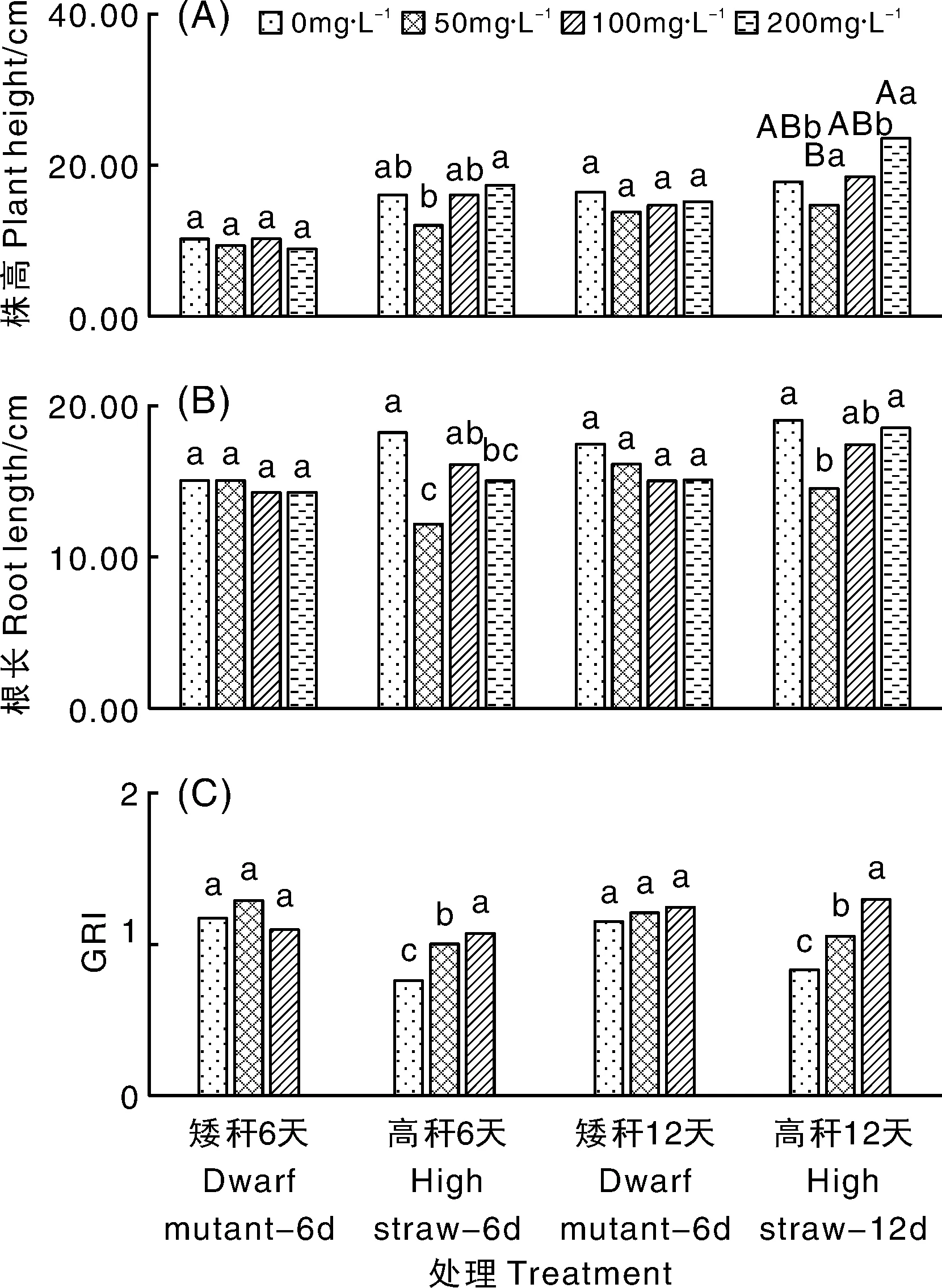
不同质量浓度处理间没有相同小写字母表示差异显著(P<0.05),没有相同大写字母表示差异极显著(P<0.01)。The uppercase letters or lowercase letters on the bars showed significantly different at the level of 0.01 or 0.05, respectively.图3 不同质量浓度赤霉素对糜子株高、根长和GRI的影响Fig.3 Effect of different concentrations of gibberellin on plant height, root length and GRI of Panicum miliaceuml
由图3-B可以看出,第6天和第12天的矮秆材料根长在赤霉素为0、50、100、200 mg·L-1不同组之间差异不显著(P>0.05)。第6天时,赤霉素0 mg·L-1组的高秆材料根长要显著高于赤霉素50 mg·L-1和赤霉素200 mg·L-1组(P<0.05),与赤霉素100 mg·L-1组之间无显著差异(P>0.05)。第12天时,赤霉素0 mg·L-1和赤霉素200 mg·L-1组的高秆材料根长要显著高于赤霉素50 mg·L-1组(P<0.05)。
由图3-C可以看出,第6天和第12天,赤霉素不同浓度组的矮秆材料GRI无显著差异(P>0.05)。第6天和第12天,高秆材料GRI在赤霉素不同浓度组之间差异显著(P<0.05),且随赤霉素浓度的递增呈现递增趋势。
3 讨论
目前,关于糜子矮秆突变体并无相关报道,但水稻中已有80多个矮化突变体被报道,根据表型性状的不同,将它们分为多种类型。本研究首次报道了糜子矮秆突变体,通过对糜子矮秆突变体778进行形态学观察以及对田间农艺性状进行测定,结果显示,矮秆突变体778的穗长和株高极显著低于高秆260(P<0.01),这与Chen等[24]在由小麦品种Sumai 3的乙基甲基磺酸盐处理产生的矮秆突变体NAUH164的研究结果相一致。矮秆突变体778的种子长也显著高于高秆,和Lu等[25]在小麦新型矮化突变体NM9中的研究结果一致。本研究揭示了矮秆突变体共有的一些典型特征,为糜子矮秆资源的利用和开发奠定了基础。
关于植株矮化的分子机制已经有很多报道。已从水稻中克隆出30多个矮化相关基因,它们大多参与赤霉素合成或转导途径。Ueguchi-Tanaka等[26]报道了一种新的GA不敏感的水稻矮秆突变体gid1,其GID1基因编码与激素敏感性脂肪酶相似的未知蛋白质,重组谷胱甘肽S-转移酶(GST)-GID1仅对生物活性GAs具有高亲和力,而对应于3个gid1等位基因的突变GST-GID1不具有GA结合亲和力。此外,GID1在酵母细胞中以GA依赖性方式与水稻DELLA蛋白SLR1结合。GID1过表达导致GA-过敏表型,这些结果表明,GID1是介导水稻GA信号传导的可溶性受体。Spielmeyer等[27]通过来自水稻基因组的序列数据与先前的作图研究相结合,以在染色体1上的sd-1的预测图谱位置定位推定的GA 20-氧化酶基因(Os20ox2),提出半矮化(sd-1)表型是由于缺陷的20-氧化酶GA生物合成酶引起的伸长节中活性GAs缺乏的结果。王翠红等[28]将丽江新团黑谷用EMS(ethyl-methane sulfonate,甲基磺酸乙酯)诱变,通过图位克隆和功能基因互补验证确认该突变体是一个新的D1基因等位突变体,其D1基因在第6外显子与内含子结合处的第2 522位碱基由G突变为A,导致第6外显子被剪切掉而无法翻译出有功能的Gα亚基。
综上所述,植株矮化与赤霉素合成或转导途径有关。对于矮秆突变体,可以通过赤霉素敏感性实验,若突变体在赤霉素处理后可以恢复至正常株高,则说明矮化突变与赤霉素合成通路有关;反之,若对赤霉素处理不敏感,则可能是赤霉素传导途径或其他途径改变导致的矮化。因此在本研究中,我们通过使用GA处理矮秆突变体,发现经过喷施赤霉素后,矮秆突变体可以恢复至正常株高,从而进一步判断其矮化原因与赤霉素合成途径有关。
