遮阴对米槁幼苗光合生理指标的影响
2019-05-08邓明明刘济明文爱华李丽霞
邓明明,刘济明,文爱华,熊 雪,李 佳,李丽霞
(贵州大学 林学院,贵州 贵阳 550025)
光能作为植物光合作用能量的主要来源,也是植物生长的关键.植物类型不同,对光强适应需求程度也不相同.不同遮阴下,植株会通过改变形态结构、光合色素含量和光合速率大小等来适应环境[1-6].在弱光条件下,植物通过降低光补偿点来提高光能利用率,以达到弱光下的最大光合速率,积累有机物.同一弱光条件下,耐阴植物的光合效率比不耐阴的高,弱光下植物光合作用也会受到一定限制.耐阴植物对光的响应与喜光植物有较大差异[7-11],主要表现在光补偿点和光饱和点更低,净光合速率较高[12-13].通常遮阴下的植株叶绿素总含量会升高,叶绿素a/b含量比值下降,使植株在弱光下的光能利用率得到提高[14];弱光时类胡萝卜素含量升高,捕光能力升高,以避免紫外辐射对植株的伤害[15-16].
米槁(CinnamomummigaoH. W. Li)是樟科(Lauraceae)樟属的常绿乔木,别名大果木姜子[17-18],是我国少数民族药材之一,其果可用于治疗腹痛、呕吐、晕车等疾病,也可作为香料、工业原料等,主要分布于广西、贵州、云南3省区交界处[19].近年来,米槁野外种群数量急剧下降,资源匮乏,药源紧张.野外调查发现,现存米槁多为成熟植株,林下幼苗较为稀少,且可见死亡迹象[20].作为群落先锋树种,其林下幼苗极有可能是因光照不足限制了生长.作为药用植物,其需求量较大,产量开始明显下降,因此米槁培育技术和生长特征开始受到关注.现阶段关于米槁的研究主要为不同氮磷、水分条件下其光合作用对CO2响应及相关生理变化,国外未见米槁相关研究报道[21-27].笔者采用人工遮阴手段,研究米槁幼苗对不同光照的光合响应特性,了解光因子对米槁幼苗生长的影响,拟为探索米槁生长培育条件提供更多科学依据.
1 材料与方法
1.1 供试材料
通过白纱布与黑尼龙网试验设置3种透光处理,其中:全光照L1:透光率100%;中度遮阴L2:80目单层白色纱布,透光率75% ± 5%;重度遮阴L3:单层黑色尼龙网,透光率40% ± 5%(各透光率通过北师大光电 ST-85型照度计在天气晴朗条件下确定).每个处理20盆,每天在17:00—18:00对花盆称量并调节水分状况(W1:土壤相对含水量80% ± 5%;电子秤感量5 g).试验开始于2015年6月25日,胁迫试验持续120 d左右,每隔40 d测量1次数据,共分前、中、后3个时期,期间不施肥并随时清除杂草.
1.2 测定方法
在试验的每个时期,每个处理分别选取相同部位3 ~ 5片成熟叶片(选取每个遮阴处理下3个重复处理测定指标),立即带回实验室利用乙醇提取法进行光合色素提取,同时测定不同透光处理下叶片叶绿素a(Chla)、叶绿素b(Chlb)、总叶绿素(Chl)以及类胡萝卜素(Car)的含量4项指标,测定方法参考文献[28].
在不同遮阴处理下的各20盆中选3盆长势相同的植株,每盆重复下择3片长势相同的叶片,共9片作为测定重复叶片(受光方向一致的上部叶片),于试验后期连续晴朗无风(或微风)天的9:00—11:30进行光响应测定.用光合作用测定系统(美国Li-cor公司生产的Li-6400)测定当天外界平均光照辐射强度,并作为额定光强.其中净光合速率Pn、气孔导度Gs、胞间CO2浓度Ci、蒸腾速率Tr等光合指标的测定,在额定光强和叶室CO2浓度为(380 ±2) μmol·mol-1条件下进行.测定各光合参数前,通过预试验得到叶片光饱和点在1 800 μmol·dm-2·s-1左右,为避免高光产生抑制,确定活化光强为1 500 μmol·m-2·s-1,光合诱导时间为5 min.对测定系统的02B-LED红蓝光源进行设定,在65% ± 5%相对空气湿度、(20 ± 0.5) ℃叶温下,测定在光合有效辐射PAR值分别为:2 000, 1 800,1 500,1 200,1 000,800,500,300,200,100,80,50,0 μmol·m-2·s-1下的4个指标Pn、Gs、Ci、Tr,每个PAR设置值停留200 ~ 150 s.通过平均值分析,根据测定数据计算WUE 值.WUE 计算公式如下:WUE =Pn/Tr[21].
值得注意的是,测定前应达到以下条件:①气体流速500 μmol·s-1;②叶室中CO2浓度控制在(380 ± 2) μmol·mol-1;③光强1 500 μmol·m-2·s-1下活化10 min.
1.3 数据统计与分析
所有数据分析通过SPSS 19.0处理,图表采用Excel 2007软件进行处理.
2 结果与分析
2.1 遮阴对米槁幼苗光合色素的影响
米槁叶片光合色素Chla、Chlb、Chl、Car含量随光照强度变化而变化的情况如图1所示.

图中所有光合色素的含量均为鲜质量下的测定值.图1 不同遮阴下米槁叶片光合色素含量的变化
由图1可知,米槁叶片光合色素Chla、Chlb、Chl、Car含量随光照强度的降低而增加,且在重度遮阴L3条件下,光合色素含量最高.以试验后期为例,分别为:2.26,1.87,4.13,0.82 mg·g-1(鲜质量),相比全光照条件下增加0.9,2.67,1.43,1.56倍,同L1、L2条件下的光合色素含量差异明显.在光合作用过程中,遮阴能提高米槁叶片光合色素含量,透光率越低光合色素含量越高,而轻度遮阴与全光照下差距不大.随着试验继续进行,Chla含量随处理时间延长而增加,但在L2、L3条件下最大值均出现在试验中期,分别为0.91 mg·g-1和2.04 mg·g-1;后期略有下降,L1处理下整体变化不大,但呈持续上升趋势.Chlb含量随着处理时间的延长,略有下降,最大值均出现在前期,分别为L1:0.57 mg·g-1,L2:0.97 mg·g-1,L3:2.12 mg·g-1.而Chl、Chla/b、Car含量皆随时间的延长而增加,3个遮阴处理下最大值均出现在试验中期,分别为Chl:1.70,2.24,4.41 mg·g-1;Chla/b:2.68,1.53,1.16 mg·g-1;Car:0.35,0.58,0.89 mg·g-1,后期均略有下降.且Chla/b含量比值在试验处理前期最低(1.96 mg·g-1),中期最高(2.68 mg·g-1),后期次之(2.42 mg·g-1).说明随着遮阴时间的延长,米槁为适应环境变化,其利用弱光的能力有所增强,在透光率过低时,利用弱光能力明显降低.
2.2 遮阴对米槁幼苗光响应的影响
不同遮阴下米槁幼苗光响应的状况如图2~5所示.
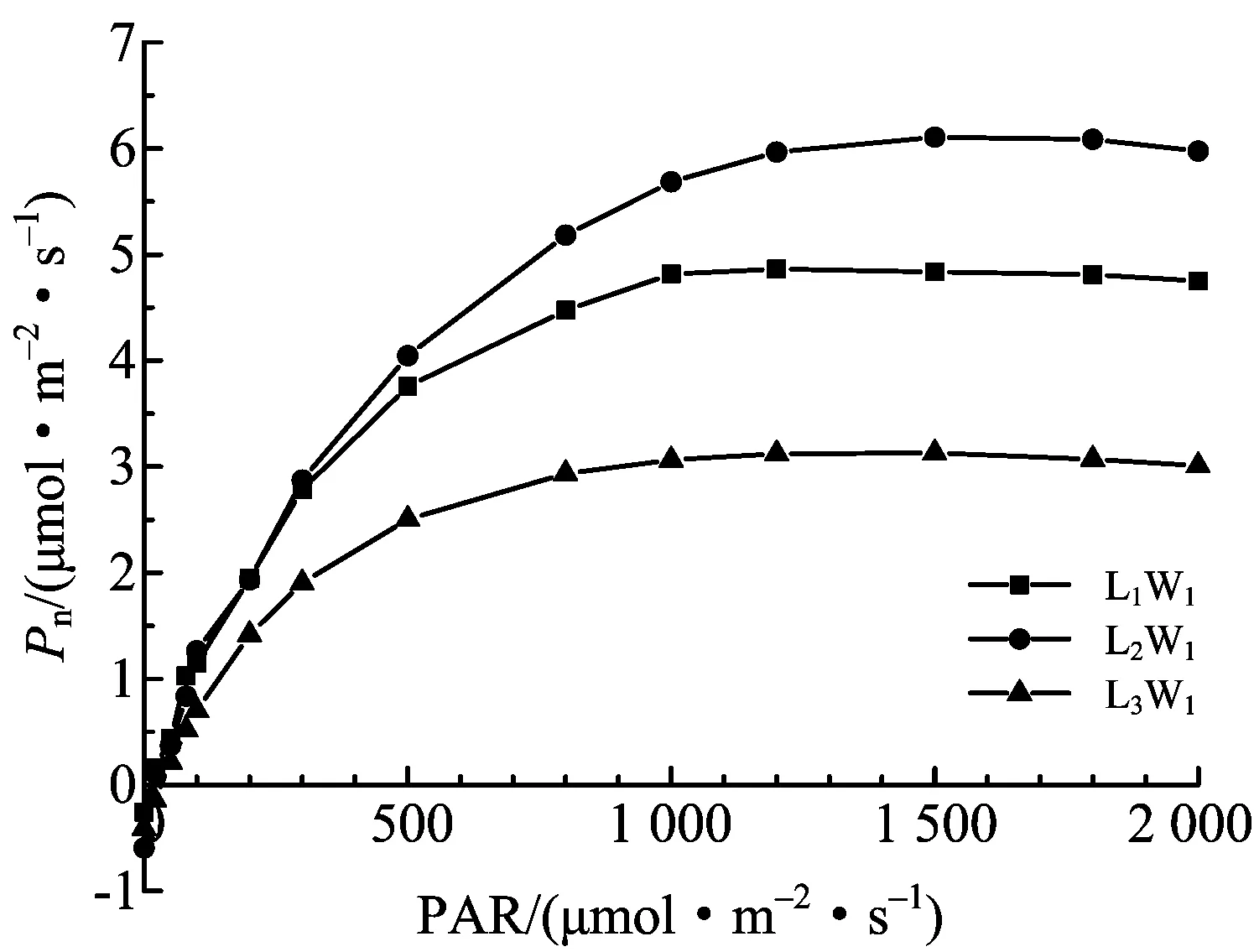
图2 净光合速率在不同遮阴下的光响应

图3 胞间CO2浓度在不同遮阴下的光响应
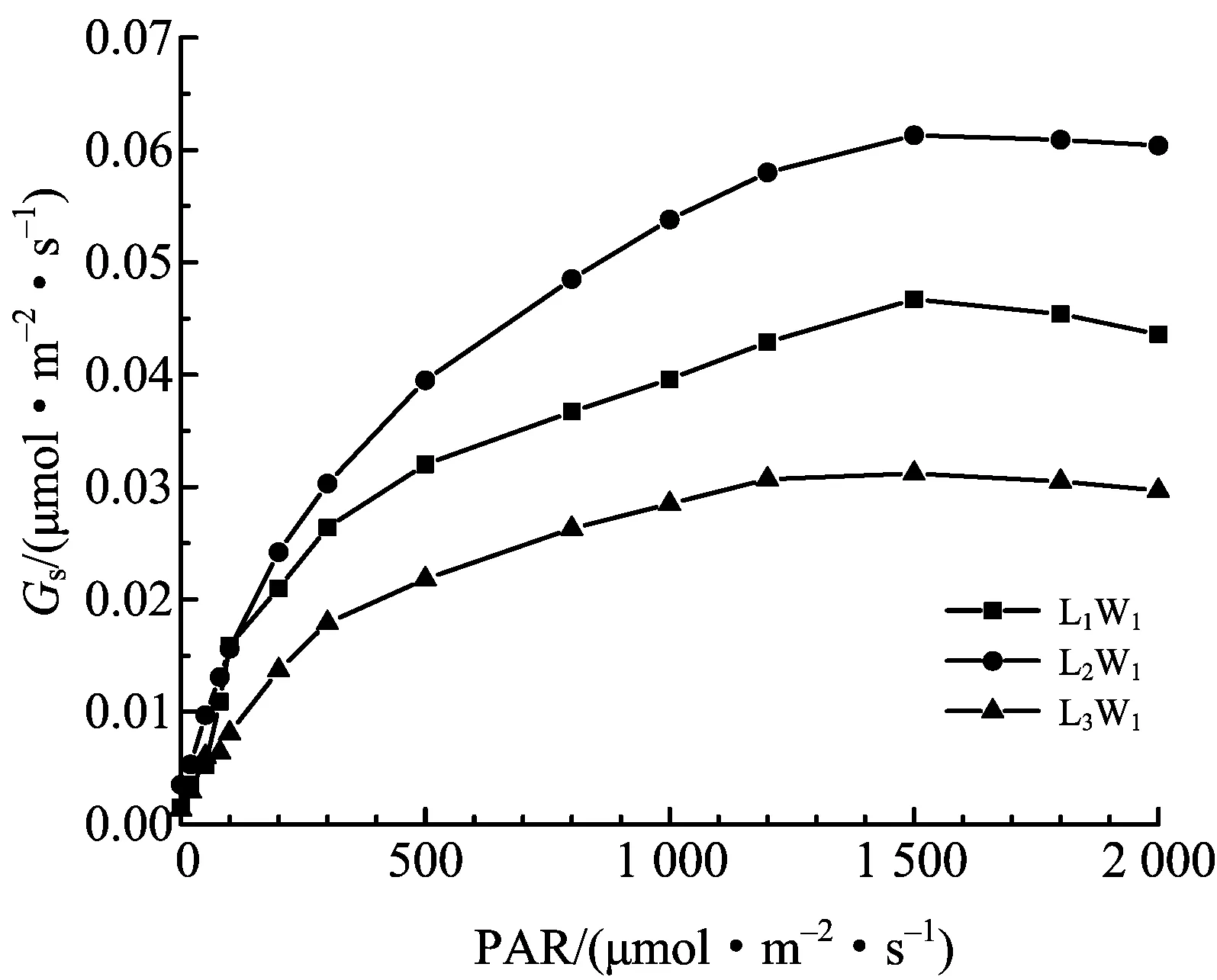
图4 气孔导度在不同遮阴下的光响应

图5 蒸腾速率在不同遮阴下的光响应
由图2可知,米槁净光合速率Pn随着PAR的升高而升高,其中在不同遮阴处理下Pn值大小均表现为L2> L1> L3,且在PAR≤100 μmol·m-2·s-1时急速上升,之后上升速度减缓,并在PAR=1 200 μmol·m-2·s-1(L1处理)、PAR=1 500 μmol·m-2·s-1(L2、L3处理下)时,达到最大,随后有所降低.胞间CO2浓度Ci随着PAR的增强而降低(图3),在PAR≤100 μmol·m-2·s-1时呈近急速下降趋势,分别在PAR=1 200 μmol·m-2·s-1(L3处理)、PAR=1 500 μmol·m-2·s-1(L1处理)、PAR=1 800 μmol·m-2·s-1(L2处理)时达到最低,随后有所回升,且3个遮阴处理下,L2处理下Ci值最低,总体表现为L2< L1< L3.与之相反,Gs(图4)、Tr(图5)皆随着PAR的增强而升高,在PAR≤100 μmol·m-2·s-1时呈近急速上升趋势,并最终趋近平缓,且分别在PAR=1 500 μmol·m-2·s-1时达到最高,随后有所降低.在各遮阴条件下,L2处理下Gs最高,表现为L2> L1> L3;L1处理下Tr值最高,表现为L1> L2> L3.
2.3 遮阴对米槁幼苗光合生理指标的影响
不同遮阴下米槁幼苗叶片光合生理指标变化趋势如图6所示.
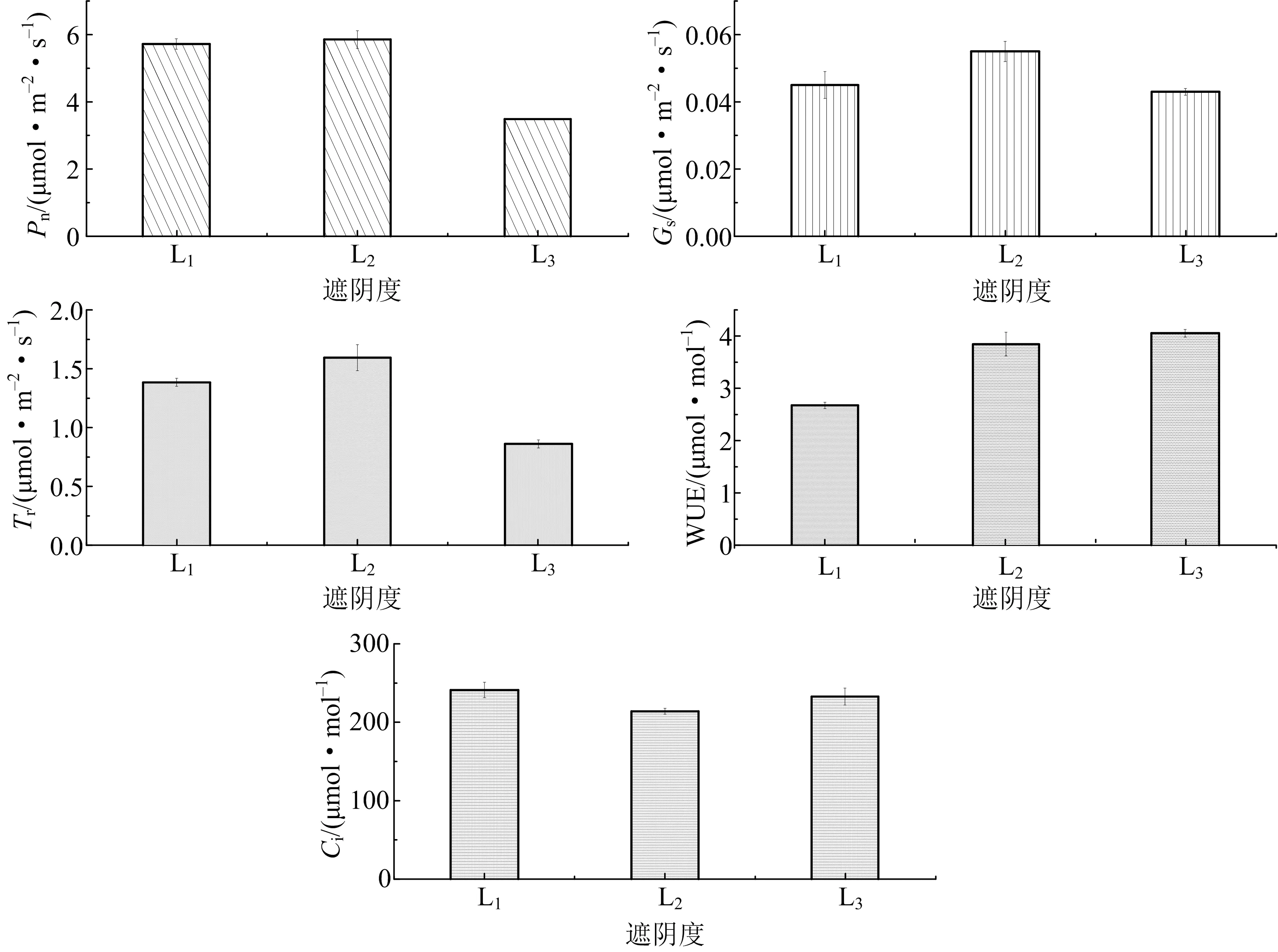
图6 不同遮阴下叶片光合生理指标变化趋势
由图6可知,在光合有效辐射强度为1 200 μmol·m-2·s-1时随着遮阴加剧,米槁Pn、Gs和Tr皆呈先增大后快速减小,L3遮阴处理下的下降受到显著抑制.与重度遮阴L3及全光照L1处理相比,在L2处理下Gs和Tr明显升高,Pn变化不显著;Ci恰好相反,随着遮阴加剧其呈现先下降后急速上升趋势,在L3处理下出现最大变化,与其他指标表现出相反的差异.而WUE则随遮阴的加剧不断上升,与L1处理相比,L3处理下WUE显著升高.
表1为米槁幼苗叶片光合作用生理指标相关性系数.

表1 米槁幼苗叶片光合作用生理指标相关性系数
注:*在 0.05 水平(单侧)上显著相关.
由表1可知,Pn同Gs呈正相关性,同Tr呈显著正相关性,同Ci、WUE呈负相关性,其中Tr与Pn相关系数最大,为0.991,说明Tr对Pn影响最大.Gs同Ci、WUE呈负相关性,同Tr呈正相关.Ci与Tr呈负相关,与WUE呈正相关.Tr与WUE呈负相关.
3 讨 论
植物光合生理变化过程是植物适应环境变化的具体表现之一,在受到环境胁迫时植物会通过改变光合色素的含量影响光合作用来适应环境变化,达到有机物的积累[29-32].通常叶绿素在光合作用中承担能量吸收转换作用,而类胡萝卜素则担任着捕光及光防御作用[33].试验中,随着透光率的降低,米槁Chla、Chlb、Chl、Car含量逐渐升高,在重度遮阴下Chla、Chlb、Chl、Car含量明显高于全光照、中度遮阴处理且差异显著,而重度遮阴下Chla/b含量比值不断下降是由于植物对弱光产生的适应反应,这同薛伟等[34]对疏叶骆驼刺、张云等[35-38]对堇叶紫金牛(Ardisiaviolacea)、美国山核桃等的研究结果一致.试验中,米槁通过增加光合色素含量来适应不良的光照环境,但是遮阴仍然导致了幼苗光合速率降低.同时有研究表明,耐阴植物Chla/b含量的比值约为2.3,喜光植物Chla/b的约为3[39-40],米槁Chla/b介于两者之间且偏向于喜光,说明米槁不是耐阴植物.
植物光补偿点及光饱和点是植物对光利用能力的体现,代表植物对光的需求特性和需求量[32].米槁的Gs、Tr、Pn值在重度遮阴下上升缓慢且低于全光照和中度遮阴,可见米槁具有较高的光饱和点,且随着透光率的改变有所不同.米槁全光照下光饱和点均低于遮阴处理,但对应于Pnmax却表现出Pnmax L2>Pnmax L1>Pnmax L3的规律,说明对米槁进行遮阴并未降低其光饱和点,但轻度遮阴能提升米槁的Pn.有研究证明,耐阴植物在遮阴处理下表现出光饱和点和光补偿点降低、植物Pn上升的现象[8,10],这与笔者研究结果相违背.阳生植物叶片光饱和点较高,光合辐射在2 000 μmol·m-2·s-1以内都能进行较好的光反应[41].但是米槁在全光照下净光合速率Pn低于中度遮阴条件,说明米槁不适于全光照下生存.
植物净光合速率Pn是了解植物光合能力的重要指标,Pn往往受到Ci、Gs、Tr的影响.气孔的开闭直接影响植物胞间水分及胞间CO2浓度,进而影响植物的蒸腾、光合作用[42],Gs能直接反映植物气孔的开张情况,并对环境改变反应敏感.米槁Gs随着遮阴程度的升高表现出先升后急速降低趋势,Pn反应的趋势类似于Gs,Ci与Gs相反,体现了两者间的反馈调节作用[43].但Gs上升时,Tr却持续下降,说明气孔并非限制因子.从相关性分析看,Pn同Tr的相关系数最高,说明蒸腾作用对光合能力影响最大.全光照下,米槁Pn、Gs值均低于轻度遮阴,说明米槁幼苗可能出现轻微强光抑制现象,导致气孔有所关闭所致[44].而适当遮阴缓减了强光对米槁的胁迫,气孔张开,从而有利于Pn的提升[45].但PnmaxL2与PnmaxL1差距不大,且随着遮阴的加剧,Chla、Chlb、Chl、Car含量持续上升,Tr持续下降,光合色素含量的改变及蒸腾作用可能是该现象产生的原因.过度遮阴处理则明显抑制了米槁光合作用,也说明米槁耐阴程度不大,其弱光利用率不高,仅有一定适应性.文爱华[26]研究发现,米槁幼苗在中度遮阴下可溶性糖、可溶性蛋白、游离脯氨酸的含量和3种抗氧化酶的活性出现最小值,叶片含水量则在同一时段中度遮阴下出现最大值.这与笔者的该研究结果一致,表明中度遮阴下米槁生长状况最好.
4 结束语
研究结果表明,米槁应属于阳生植物,具有较高光饱和点,且对弱光具有一定适应性,透光率在75% ± 5%下有利于提高米槁光合作用能力,在培育米槁时可适量遮阴.但米槁耐阴能力有限,在透光率40% ± 5%条件下,光合能力明显降低,但米槁的正常生长及光合作用能力都说明,遮阴还在承受范围内,其耐阴极限仍有待进一步探寻.米槁的致危因子还需进一步探索,可能与植物化感作用或本身群落演替更新有关.
