过表达血红素加氧酶-1对低氧训练小鼠骨骼肌纤维组成的影响※
2019-05-08司立宁秦海兰甘罗曼赵延礼
司立宁,王 嵘,秦海兰,崔 浩,甘罗曼,赵延礼*
(1.青海大学附属医院,青海 西宁 810001;2.青海大学基础医学研究中心,青海 西宁 810016)
低氧训练已被广泛应用于各类体育项目,被认为是提高有氧耐力成绩的良好辅助练习方法,目前就其机制研究主要集中在血液指标及最大吸氧量、毛细血管数目等方面。本研究主要关注血红素加氧酶-1(Heme Oxygenase-1,HO-1)在低氧训练中发挥的作用及其对骨骼肌纤维组成的影响。
1 材料与方法
1.1 主要试剂和仪器选择
SPF级雄性过表达HO-1转基因小鼠和野生小鼠购自赛业生物科技公司。Trizol试剂、逆转录试剂盒、Real-time PCR试剂盒均购自Invitrogen公司;Myoglobin和PGC-1α蛋白兔抗多克隆抗体购自Abcam公司;山羊抗兔IgG抗体购自北京中杉金桥公司。小动物跑台购自泰盟科技有限公司;7500型号Realtime-PCR仪购自ABI公司。
1.2 动物分组及训练
将SPF级雄性过表达HO-1转基因小鼠和野生小鼠饲养8 w后进行实验。小鼠分为6组(每组15只),即野生小鼠低氧静息组(C)、野生小鼠低氧训练30d组(C+T30)、野生小鼠低氧训练60d组(C+T60)、过表达HO-1转基因小鼠低氧静息组(H)、过表达HO-1转基因小鼠低氧训练30d组(H+T30)和过表达HO-1转基因小鼠低氧训练60d组(H+T60)。
小鼠饲养和训练在青海大学基础医学研究中心(海拔2261m,氧含量16.3%)进行。C组和H组小鼠仅做适应跑台运动训练,C+T30组、C+T60组、H+T30组和H+T60组小鼠除做适应跑台运动训练外,还需做低氧训练(20m/min,坡度5°,60min/d,5d/w,30d和60d)。各组小鼠均于末次训练结束3 d后予持续运动(20m/min,坡度5°),直至小鼠伏地,毛刷、声、光、电刺激尾部均无效或人为驱赶都无法使小鼠继续运动视为力竭,记录力竭运动时间,力竭运动后即刻采用直接颈椎脱臼法处死小鼠,快速分离双侧腓肠肌。
1.3 红肌纤维、白肌纤维面积百分率计算
肌纤维切片行ATP酶染色,Ⅱ型肌纤维显深棕色或黑色,Ⅰ型肌纤维显浅棕色或不显色。切片封固后,采用Simple PCI显微图像分析仪拍照,每张切片选取5个独立区域,利用Image pro plus 5.1系统进行图像分析,计算红肌纤维和白肌纤维的面积百分率。
1.4 real time PCR检测
采用Trizol试剂提取各组小鼠腓肠肌组织总RNA。取总RNA 2 μg逆转录合成第一链cDNA。以第一链cDNA为模板,以realtime PCR法扩增myoglobin,内对照为GAPDH。
1.5 免疫印迹检测
每20 mg组织加入250 μL 含PMSF的蛋白裂解液,在冰上充分匀浆后离心(4℃,13500r/min,15min),吸取上清液于管中,取少量蛋白样品测定浓度,其余蛋白样中加入上样缓冲液,于沸水中煮10 min,行十二烷基磺酸钠/聚丙烯酰胺凝胶电泳后转移至PVDF膜,用5%脱脂牛奶封闭后孵育第1、2抗体后行ECL显色曝光。
1.6 统计学处理
全部实验数据采用SPSS13.0数据统计软件进行处理,结果用均数±标准差表示。组间比较采用方差分析,多重两两比较采用LSD检验,以P<0.05为差异有统计学意义。
2 结果
2.1 各组小鼠力竭运动时间(表1)
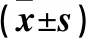
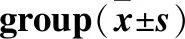
Table 1Exhaustive exercise time of mice in each
*:与C组比较P<0.05;▲:与H组比较P<0.01;#:与C+T30组比较P<0.05;⊕:与C+T60组比较P<0.01
与C组相比,C+T30、C+T60组小鼠力竭运动时间增加(P<0.05);与H组相比,H+T30、H+T60组小鼠力竭运动时间显著增加(P<0.05),并且与低氧训练野生小鼠相比,过表达HO-1转基因小鼠低氧训练30 d和60 d后的小鼠力竭运动时间显著增加(P<0.05)。
2.2 各组小鼠腓肠肌纤维横断面积(表2)



分组n红肌纤维白肌纤维C15 15.28±3.73 47.33±2.22H15 16.47±1.68 46.19±2.59C+T3011 16.69±1.78 46.37±2.14H+T3012 19.81±3.48▲# 45.25±2.72C+T6011 18.24±2.66∗ 44.91±3.63H+T609 27.42±2.74▲⊕ 41.38±3.68▲F- 25.36 5.64P- 0.00 0.00
*:与C组比较P<0.05;▲:与H组比较P<0.01;#:与C+T30组比较P<0.05;⊕:与C+T60组比较P<0.01
与C组相比,C+T30组小鼠红肌纤维面积略有增多,C+T60组小鼠红肌纤维面积显著增多(P<0.05);与H组相比,H+T30、H+T60组小鼠红肌纤维面积增多(P<0.01),H+T60组小鼠白肌纤维减少(P<0.01),并且与低氧训练的野生小鼠相比,H+T30、H+T60组小鼠红肌纤维面积增加更显著(P<0.05),H+T60组小鼠白肌纤维减少(P<0.01);另外,与H+T30组相比,H+T60组小鼠红肌纤维面积增加显著(P<0.01),H+T60组小鼠白肌纤维减少(P<0.01)。
2.3 各组小鼠腓肠肌中myoglobin的表达情况(表3和图1)

分组nRelative Myoglobin mRNA levelRelative Myoglobin protein levelC15 1.00±0.12 1.00±0.25H15 1.27±0.08 1.16±0.17C+T3011 1.33±0.25∗ 1.48±0.18∗H+T3012 2.64±0.18▲# 2.25±0.24▲#C+T6011 1.77±0.22∗ 1.18±0.23∗H+T609 4.19±0.28▲⊕ 5.14±0.15▲⊕F- 425.04 1222.88P- 0.00 0.00
*:与C组比较P<0.05;▲:与H组比较P<0.01;#:与C+T30组比较P<0.05;⊕:与C+T60组比较P<0.01

图1 各组小鼠腓肠肌中Myoglobin的表达图
与C组相比,C+T30、C+T60组小鼠腓肠肌中myoglobin mRNA的表达增加;与H组相比,H+T30、H+T60组小鼠腓肠肌中myoglobin mRNA的表达显著增加(P<0.05),并且与低氧训练野生小鼠相比,H+T30、H+T60组小鼠腓肠肌中myoglobin mRNA的表达显著增加(P<0.05)。Western blot结果显示,与C组相比,C+T30组小鼠腓肠肌中myoglobin蛋白的表达增加(P<0.01);与H组相比,H+T30、H+T60组小鼠腓肠肌中myoglobin蛋白的表达显著增加(P<0.01),并且与低氧训练野生小鼠相比,H+T30、H+T60组小鼠腓肠肌中myoglobin蛋白的表达显著增加(P<0.05)。
2.4 各组小鼠腓肠肌中PGC-1α的表达情况(图2和表4)

图2各组小鼠腓肠肌中PGC-1α的表达图
Figure2The expression of PGC-1α in gastrocnemius muscle of mice in each group
Table 4The Relative protein expression of PGC-1α in gastrocnemius muscle in each
*:与C组比较P<0.05;▲:与H组比较P<0.01;#:与C+T30组比较P<0.05;⊕:与C+T60组比较P<0.01
与C组相比,C+T30、C+T60组小鼠腓肠肌中PGC-1α的表达均显著增加(P<0.05);与H组相比,H+T30、H+T60组小鼠PGC-1α的表达均显著增加(P<0.05),并且与低氧训练野生小鼠相比,H+T30、H+T60组小鼠腓肠肌中PGC-1α的表达显著增加(P<0.05)。
3 讨论
本研究发现低氧训练30 d或60 d可延长野生小鼠力竭运动时间,而且低氧训练60 d可引起骨骼肌红肌纤维增多和myoglobin、PGC-1α的表达增加;过表达HO-1能够进一步延长小鼠力竭运动时间,并且低氧训练30 d和60 d都可引起骨骼肌红肌纤维增多和myoglobin 、PGC-1α的表达增加。
低氧训练是在运动训练周期中持续或间断采用低氧条件刺激,利用高原自然低氧环境或人工模拟低氧对人体所产生的特殊生物学效应,有利于提高运动中的抗缺氧生理反应及适应能力,进而达到提高运动成绩的目的[1]。30多年来的研究证实低氧训练可通过多种途径达到提高运动耐力的目的[2-4]。常凤等研究发现一定范围内高表达HO-1能够延长小鼠力竭运动时间,促进运动性疲劳的恢复[5]。本研究发现,野生小鼠经30 d或60 d低氧训练后力竭运动时间延长,运动耐力增加。本研究获得的另一个重要发现是,过表达HO-1的小鼠经30 d或60 d低氧训练后运动耐力增加更加显著,提示HO-1在低氧训练中具有延长小鼠力竭运动时间、提高运动耐力的作用。
骨骼肌通常可分为红肌纤维(氧化型慢肌)和白肌纤维(糖酵解型快肌)。红肌纤维富含线粒体,依赖氧化代谢产能,收缩速度慢,不易疲劳,富含myoglobin,呈暗红色。白肌纤维线粒体数目较少,依赖糖酵解途径供能,收缩速度快,易疲劳,呈淡红色[6]。有关低氧训练对骨骼肌肌纤维的影响存在两种不同的观点,一种观点认为低氧训练可以促进骨骼肌红肌纤维增加;另一种观点认为低氧训练不影响骨骼肌肌纤维类型的转变。Bigard[6]发现Wistar 鼠在低压氧舱(5500m)行低氧训练4 w后观察到红肌纤维增加[7]。Hoppeler 等发现运动员低住高训(3850m)6 w后红肌纤维富含的myoglobin 蛋白表达显著升高[8]。相反,Perhonen对于Wistar鼠行低氧(2500m)训练后的肌纤维组成无明显变化[9]。Wistar鼠行低氧(4000m)游泳训练14 w的研究显示腓肠肌纤维分布不受低氧和训练的影响[10]。我们在研究中发现,野生小鼠在低氧训练60 d后腓肠肌中红肌纤维增多,过表达HO-1小鼠在低氧训练30 d和60 d时都引起了红肌纤维显著增多,与此同时都伴有myoglobin的表达增加,提示HO-1在低氧训练中具有促进骨骼肌红肌纤维增多的作用。由此看出,不同形式的低氧训练、不同的海拔高度、不同的训练时间以及不同动物遗传背景都会导致研究结果的差异,因此对于低氧训练影响肌纤维组成的结论有待进一步验证。
PGC-1α是过氧化物酶体增殖物激活受体γ辅激活因子1,在调节肌纤维类型的转化和线粒体生物合成中发挥重要作用。PGC-1α在骨骼肌中的不同表达水平对运动能力产生很大影响。骨骼肌特异过表达PGC-1α促进肌纤维从白肌纤维向红肌纤维转化[11]。相反,骨骼肌特异性敲除PGC-1α可引起红肌纤维向白肌纤维转化,并且相对于野生小鼠,骨骼肌特异性敲除PGC-1α的小鼠力竭运动时间缩短,运动耐力下降[12]。本研究发现,野生小鼠低氧训练30 d后骨骼肌中PGC-1α的表达显著增加,并且过表达HO-1可进一步促进PGC-1α的表达增加。该结果提示HO-1在低氧训练中促进红肌纤维增加可能是通过调节PGC-1α的表达实现的。
综上所述,本研究发现过表达HO-1在低氧训练中可显著提高小鼠运动耐力,其机制可能是HO-1通过增加PGC-1α的表达促进骨骼肌红肌纤维增加实现的。
