斑皮桉5个种源光响应特性比较分析
2019-04-26唐庆兰蒋钰渝杨中宁覃明眺苏福聪邓紫宇项东云陈健波郑永邓
唐庆兰,蒋钰渝,杨中宁,覃明眺,苏福聪,邓紫宇,项东云,陈健波,郑永邓
(1.广西壮族自治区林业科学研究院 广西优良用材林资源培育重点实验室国家林业局中南速生材繁育重点实验室,南宁 530002;2.广西国有七坡林场,南宁 530227;3.环江毛南族自治县川山镇林业工作站,广西河池 547100;4.广西国有三门江林场,柳州 545000)
斑皮桉(Corymbia maculata),高大乔木,在良好生长环境下树高可达35 m,胸径达100~130 cm。分布于昆士兰州南部和新南威尔士州中部和北部沿海区域[1-2]。斑皮桉成熟材的边心材明显,边材一般呈淡白色,宽度可达8 cm,心材为浅棕色至深棕色,纹理通常交错,材质坚硬、结实、耐用。成熟材的基本密度为745~1 080 kg/m3,可用作枕木、工具柄手、重建筑、横梁、木地板、胶合板等[3]。斑皮桉是澳大利亚东部沿海重要的用材树种之一,该树种的育种和改良工作自1995年开始[4-6],建立了世代种子园,建立了多个无性系测定试验林。斑皮桉无性系林木生长性状和木材性状受较高遗传控制,环境因子对生长性状和木材材性的影响达到显著水平,遗传与环境交互作用对木材材性的影响很小,对生长性状有一定的影响[7]。我国从上世纪90年代开始进行斑皮桉引种试验[8-10],在福建,斑皮桉耐寒能力中等,生长不及尾巨桉(Eucalyptus urophylla×E.grandis)、巨尾桉(E.grandis×E.urophylla)和邓恩桉(E.dunnii);在海南,参试的无性系中,斑皮桉无性系生长量仅次于尾细桉(E.urophylla×E.tereticornis)和尾赤桉(E.urophylla×E.camaldulensis)2个杂交种无性系,高于其他无性系生长量。华南农业大学引进斑皮桉等4个伞房属树种,并开展了遗传改良研究[11-12],研究发现,引种进的4个伞房属树种中,斑皮桉的生长表现较差,3年生时平均材积0.002 m3。
桉树不同种源、不同无性系的各项光合生理指标差异显著[13-16]。陆钊华等[13]研究发现,净光合速率、气孔导度与蒸腾作用与树高、胸径、单株材积等生长指标存在一定程度的正相关关系,可应用于优良无性系的早期选择。对斑皮桉光合作用的研究未见报道,本文分析了5个种源斑皮桉的光响应特性,为斑皮桉的经营和早期选择提供参考。
1 材料与方法
1.1 试验地自然概况
试验地设在广西林科院试验苗圃(108°56′E,22°56′N),海拔约80 m,属亚热带季风气候,≥10℃积温为7 200℃,年均气温21.7℃,极端最低气温-1.5℃,极端最高气温40.4℃,全年无霜期达360 d,年均降水量为1 300 mm,日照时数1 550 h以上,年均相对湿度为80%左右,干湿季节明显,降雨主要集中在4—9月。
1.2 参试种源
2013年12月从澳大利亚联邦科学与工业研究组织引进斑皮桉,包括5个种源,主要产自澳大利亚新南威尔士州和维多利亚州(表1)。

表1 斑皮桉种源地概况Tab.1 Origin information of Corymbia maculata provenances
1.3 光合测定
2016年1月播种,小苗培育至30 cm后,将小苗移栽至直径30 cm的塑料杯内培育,2016年10月进行光合测定。
用美国LI-COR公司生产的LI-COR6400光合作用仪进行测定,每个种源选取3株平均样木,测定时间选择晴天的上午9—11时进行。测定参数包括净光合速率(Pn)、气孔导度(Gs)、胞间二氧化碳浓度(Ci)、蒸腾速率(Tr),并计算水分利用效率(WUE=Pn/Tr),光强(PAR)梯度分别设定为 2 000、1 800、1 600、1 400、1 200、1 000、800、 600、 400、 200、 100、 500、 200、 100、50、0。
1.4 数据处理及分析
本文所有数据采用Excel 2007进行数据处理和作图。
2 结果与分析
2.1 不同种源净光合速率光响应特征
各种源间光响应曲线的变化趋势一致,但又有所差异。光强为零时,净光合速率(Pn)为负值,随着光强的增加,Pn迅速上升,光强达到800 μmol/(m2·s),曲线上升趋向平稳,达到饱和点后Pn上升缓慢,甚至有下降的趋势。种源20772的Pn最大值 (22.528 4 μmol·m-2·s-1) 最大,最小值偏小,曲线上升相对较急;其次是种源20599、20884和20598,Pn最大值分别为20.232 0、18.686 4、17.817 2 μmol/(m2·s);Pn最小的是种源21075,Pn最大值为15.546 3 μmol/(m2·s),最小值偏小,曲线上升平缓(图1)。
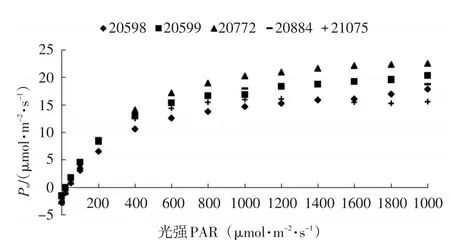
图1 不同种源Pn光响应曲线Fig.1 Pn-light response curves of different provenances
从净光合速率(Pn)与光强的回归(表2)可以看出,Pn与光强呈显著的二次曲线相关关系,各种源拟合曲线的R2值均在0.92以上。各种源的光响应曲线的二次回归拟合和200 μmol/(m2·s) 以下的线性拟合,可以计算出一系列光响应特征参数(表3),光补偿点较高的种源有20772和20598,光补偿点分别为36.92、45.05 μmol/(m2·s),但光饱和点较高,均超过1 500 μmol/(m2·s),利用强光的能力较强;种源20599和21075的光补偿点次之,分别为21.2 、21.67 μmol/(m2·s),对弱光的利用能力较强,光饱和点中等,均为 1 388.89 μmol/(m2·s),利用强光的能力中等;种源20844光补偿点最低(1.28 μmol·m-2·s-1),利用弱光的能力最强,但光饱和点较低(1 350 μmol·m-2·s-1),利用强光的能力弱。
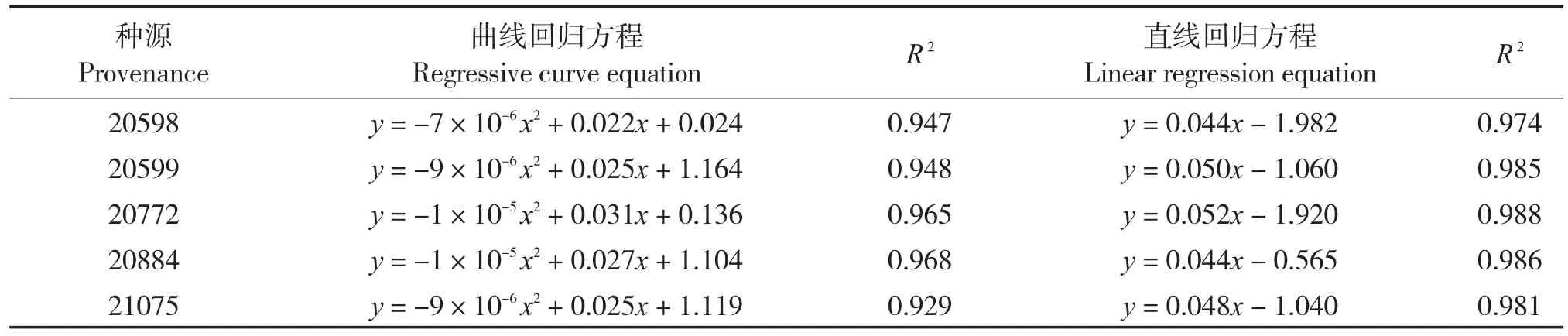
表2 二项式回归方程和光强在200 μmol/(m2·s)以下的直线回归方程Tab.2 Binomial regression equation and linear regression equation of PAR under 200 μmol/(m2·s)
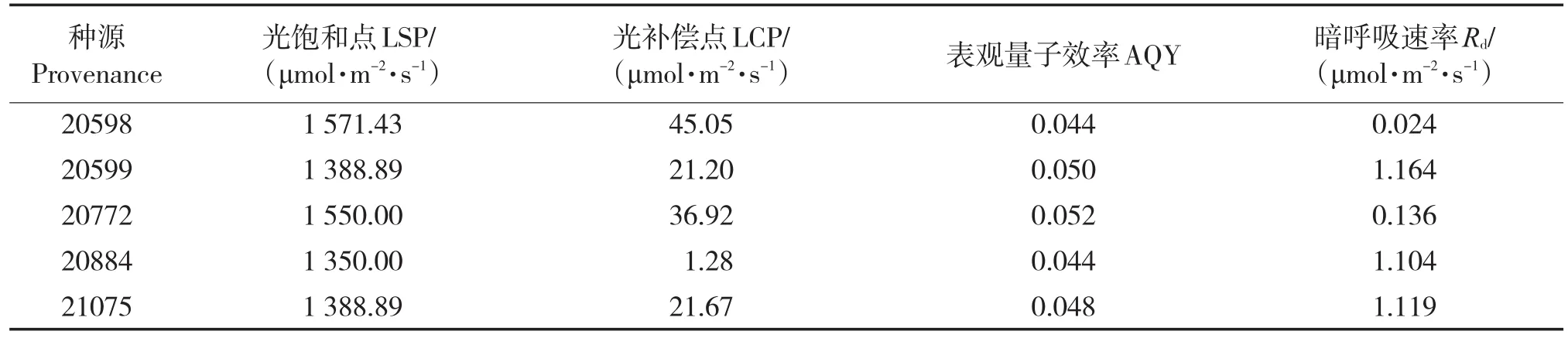
表3 5个种源斑皮桉光响应特征参数Tab.3 Light response curve parameters of 5 provenances
2.2 不同种源斑皮桉气孔导度和胞间二氧化碳的光响应
5个斑皮桉种源的气孔导度(Gs)随光强的增强而缓慢上升,各种源变化趋势一致,但种源间的变化幅度存在差异(图2)。种源20599的Gs随光梯度的变化幅度为0.078~0.430 molH2O/(m2·s),变化幅度最大;种源20772的变化幅度次之,0.134~0.364 molH2O/(m2·s),种源20598、20884和21075的变化幅度较小,0.061~0.269 molH2O/(m2·s)。

图2 不同种源气孔导度光响应曲线Fig.2 Gs-light response curves of different provenances
胞间二氧化碳(Ci)浓度随光强的增大而下降,5个种源的下降趋势基本一致,各种源间差异较小 (图3)。光强低于400 μmol/(m2·s) 时,各种源Ci浓度随光强的增加下降趋势明显,光强在400~800 μmol/(m2·s) 之间Ci浓度下降缓慢,当光强大于1 000 μmol/(m2·s)时,各种源的Ci浓度又开始缓慢上升,上升幅度比较小。

图3 不同种源胞间二氧化碳光响应曲线Fig.3 Ci-light response curves of different provenances
2.3 不同种源斑皮桉蒸腾速率和水分利用效率的光响应
不同种源的斑皮桉蒸腾速率(Tr)随光强的增强而增大,但增幅比较小,各种源曲线变化趋势一致,与气孔导度(Gs)随光强变化的趋势相似。种源20599的变化幅度最大,1.650~6.363 mmolH2O/(m2·s);种源20598和20772变化范围相近,在1.248~4.324 mmolH2O/(m2·s)之间;种源20884和21075的变化幅度最小,0.229~2.136 mmolH2O/(m2·s)(图4)。
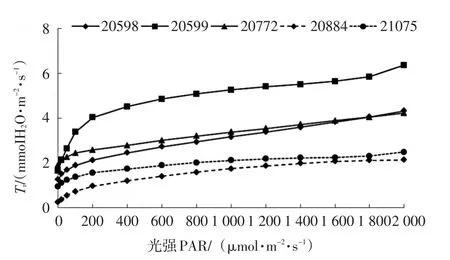
图4 不同种源斑皮桉蒸腾速率光响应曲线Fig.4 Tr-light response curves of different provenances
斑皮桉各种源水分利用效率(WUE)随光强增强而增大,各种源变化趋势基本一致,且与净光合速率(Pn)随光强变化的趋势相似。各种源在光强低于400 μmol/(m2·s)时,WUE随光强的增加迅速增加,当光强为 400~600 μmol/(m2·s)时,WUE达到最大值,随后随光强的增加而趋向平缓,甚至下降(图5)。各种源间WUE存在差异,种源20884的变化幅度最大(-4.837~10.717 molCO2/mmolH2O);种源21075的变化范围次之,(-1.756~7.679 molCO2/mmolH2O);种源20772的变化幅度为-1.331~5.991 molCO2/mmolH2O;种源50598的变化范围为-2.180~4.698 molCO2/mmolH2O;种源20599的变化范围最小(-0.976~3.409 molCO2/mmolH2O)。
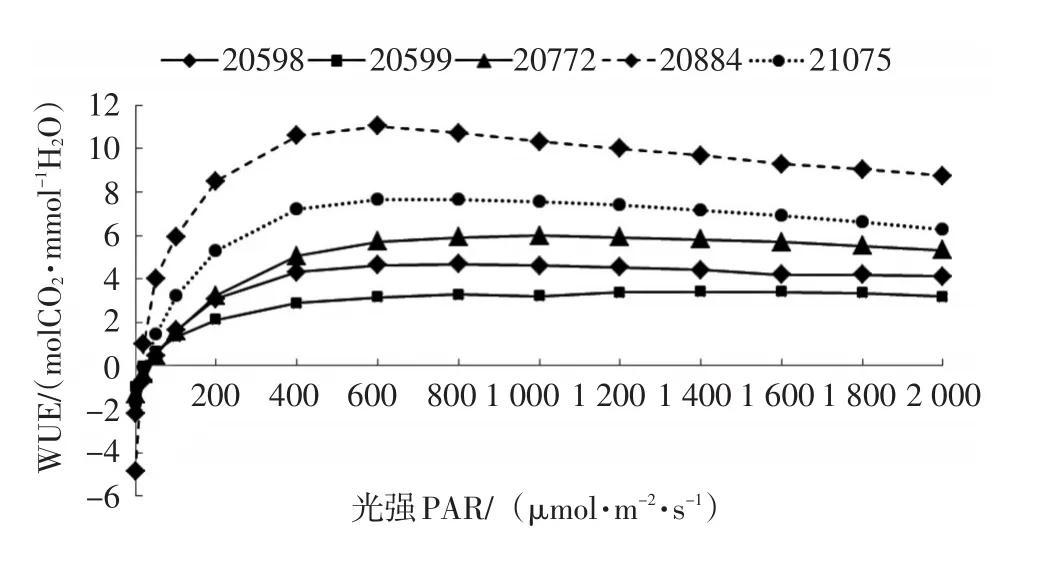
图5 不同种源斑皮桉水分利用效率光响应曲线Fig.5 WUE-light response curves of different provenances
3 结论与讨论
林木光合能力主要取决于其本身的遗传特性,和林木生长的环境也密切相关[17-18],同一树种生长在不同的环境,光合特性有所不同。通过分析不同种源斑皮桉的光合特性,可为斑皮桉的引种、改良和栽培提供依据。光饱和点和光补偿点能直接反应植物对光环境的适应能力,高饱和点、低补偿点的植物对光环境的适应能力最强,低饱和点、高补偿点的植物对光环境的适应能力较差,表观量子AQY的大小反应植物利用光能的高低[19]。
本文研究结果表明,5个种源斑皮桉的光补偿点均超过1 300 μmol/(m2·s),与桉属的巨桉(E.grandis)、尾叶桉(E.urophylla)、巨尾桉和尾巨桉等多个无性系及同为伞房属的斑皮柠檬桉的光饱和点相似,表现出明显的阳性特征[14-16]。本研究的5个种源中,种源20772和20598的光饱和点和光补偿点相对较高,在阳光充足的阳坡种植更能发挥其生长潜能;种源20599和21075的光饱和点相同,光补偿点相差不大,在5个种源中均处于中等位置,在半阳坡种植更能适应当地的光环境;种源20884光饱和点最低,适应阴坡或半阴坡地域光环境的能力较其他种源强。
由于研究时间有限,未将斑皮桉各种源的光合能力与生长量进行综合分析,但前人的研究结果表明,同一树种不同种源表现出不同的光合特性,是由于树种长期适应当地的地理、气候因子表现出的特性,这些特性可以遗传给下一代[20-21],因此在进行引种时,选择高净光合速率(Pn)的种源,能更有效的利用光能将化学能转换成有机物质,有望获得更高的生长量。
