咖啡因在高氧诱导早产仔鼠肺损伤中的修复作用研究*
2019-04-25宋亚楠吴凯峰刘晓丽郑兴惠
邓 萌,宋亚楠,程 宇,李 青,张 翔,王 欢,吴凯峰,刘晓丽,郑兴惠,黄 波△
(1.遵义医科大学第三附属医院PICU,贵州遵义 563000;2.遵义医科大学研究生院,贵州遵义 563000)
早产儿出生时肺部发育不成熟,常需要氧气支持和机械通气治疗。然而,长期暴露在高氧环境极易造成氧化应激,形成肺纤维化,使肺部发育迟缓,最终导致支气管肺发育不良(BPD)[1]。细胞外信号调节蛋白激酶(ERK)是丝裂原活化蛋白激酶(MAPKs)家族中重要成员之一,参与基因表达,细胞增殖、凋亡等重要生命过程[2]。大量研究显示,高氧条件下可以触发ERK途径[3-5]。咖啡因通常用于治疗早产儿的呼吸暂停[6-7],一项回顾性研究发现咖啡因在预防支气管肺发育不良(BPD)中有非常重要的功效[8]。然而,咖啡因的肺保护作用机制目前尚不清楚。本研究以早产 Wistar 大鼠为研究对象,复制高氧肺损伤动物模型,观察咖啡因干预后肺纤维化情况, ERK的分布及蛋白表达,并分析咖啡因干预对早产鼠高氧肺损伤修复的作用及其与MAPKs/ERK信号通路的关系,以期为临床指导用药及综合治疗BPD提供重要的临床及理论依据。
1 材料与方法
1.1材料
1.1.1实验动物 健康成年Wistar大鼠[扬州大学比较医学中心,许可证号:SCXK(苏)2012-0004]32只,其中雄性8只,雌性24只,体质量200~250 g。分为8组,每组雌鼠和雄鼠按3∶1合笼交配,第2天上午8:00查见阴栓记为妊娠第1天,将孕鼠分笼并于妊娠第19天时行剖宫产取出早产鼠。
1.1.2主要试剂 牛血清清蛋白(BSA,德国Roche公司);抗兔/鼠通用型免疫组织化学试剂盒[Real EnvisionTMHRP (rabbit/mouse),丹麦Dako公司];Phospho-p44/42(pErk)兔多克隆抗体(美国Abcam公司)。
1.2方法
1.2.1实验动物及分组 8窝(72只)早产大鼠出生12~24 h内聚拢,分配给代母鼠并分为4组,即空气+9 g/L盐水组(A+N组)、空气+咖啡因组(A+C组)、高氧+9 g/L盐水组(H+N组)、高氧+咖啡因组(H+C组),每组18只。
1.2.2高氧模型复制 H+N组和H+C组持续暴露于高氧中,A+C组和H+C组予腹腔注射咖啡因29 mg·kg-1·d-1,A+N和H+N组每天腹腔注射同等体积的9 g/L盐水。代母鼠每24小时在4组之间更换1次,以避免氧中毒,同时排除不同组间代母鼠影响。高氧暴露在自制氧箱内完成,氧流量5 L/min,连续检测氧浓度(CYS-1数字测氧仪,上海嘉定电子仪器厂),使氧浓度维持于60%~70%。早产大鼠高氧、空气暴露14 d,室温维持在24~25 ℃ ,昼夜各12 h,自由进食进水,每天开箱15 min进行清扫,无水氯化钙和生石灰吸收水蒸气和二氧化碳并每天更换。
1.2.3肺组织收集与制备 于早产仔鼠出生后3、7、14 d,各组选取6只进行取材,3.5%水合氯醛(10 mL/kg)腹腔麻醉大鼠,开胸并结扎右肺,剪开左心耳,右心室注入含肝素的生理盐水,直到左心耳流出的液体清亮为止,分离心、肺。
1.2.4肺组织指标检测 留取左中肺组织,经4%多聚甲醛固定,常规进行切片、HE染色,光镜下观察肺组织形态学改变。在100倍光镜下进行辐射状肺泡计数(RAC),每张切片计数5次,取平均值。肺湿重/干重(W/D)测定:右肺取部分组织称湿重后置80 ℃温箱,48 h后称肺干重,计算肺W/D值。
1.2.5各组肺组织胶原纤维含量检测 采用Masson染色法,NikonDSRil偏振光显微镜下观察4组仔鼠各时间点的胶原纤维分布情况。每张切片选取5个视野,采用病理图文分析系统测试阳性细胞倍数(阳性细胞倍数=阳性细胞数/总细胞数×100%)。
1.2.6免疫组织化学检测磷酸化ERK(p-ERK)在肺组织中的分布 脱蜡,水化组织切片,3%H2O2孵育5~10 min,将切片浸于0.01 mmol/L枸橼酸盐缓冲液热修复抗原,5%BSA封闭20 min,滴加pERK抗体(1∶250),4 ℃过夜。滴加辣根过氧化物酶(HRP)标记山羊抗兔免疫球蛋白G(IgG,1∶1 000),37 ℃ 孵育20 min。二氨基联苯胺(DAB)显色,苏木素轻度复染,脱水,透明,封片。结果判定:胞核和(或)胞质呈现棕黄色即为阳性细胞。
1.2.7Western blot检测肺组织p-ERK蛋白表达水平 各组分别提取肺组织总蛋白,用BCA法进行蛋白质定量,再采用Western blot方法检测肺组织p-ERK蛋白表达水平,经发光、曝光、显影后分析结果,内参蛋白为β-actin,以蛋白灰度值进行统计分析。

2 结 果
2.1各组肺组织病理学观察 A+N组、A+C组3 d时肺泡结构不规则,肺泡腔小,肺泡间隔较厚;H+N组、H+C组3 d时可见肺泡内少量红细胞及液体渗出,偶见炎性细胞和肺水肿表现。A+N组、A+C组7 d时肺泡结构逐渐清楚,无液体和炎性渗出,肺泡间隔变薄。H+N组、H+C组7 d时肺泡壁增厚,肺组织结构紊乱,肺间质细胞增加,肺泡内大量红细胞及液体渗出,炎性细胞增多。A+N组、A+C组14 d时肺泡结构逐渐清楚,无液体和炎性渗出,肺泡间隔变薄。H+N组、H+C组14 d时炎性渗出减少,肺间质增厚并伴随纤维化,见图1。
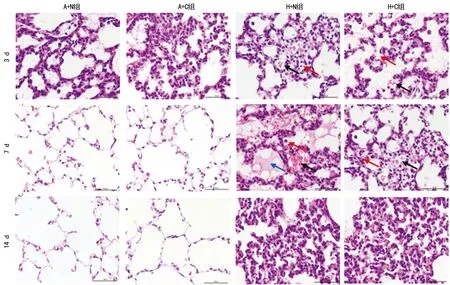
图1 各组3、7、14 d时肺组织病理结构(HE染色,×200)
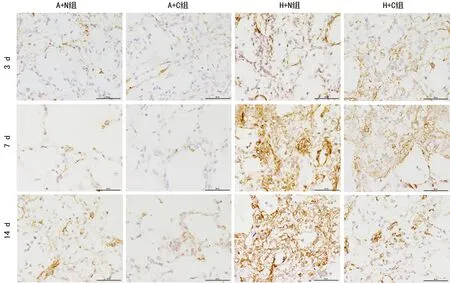
图2 各组3、7、14 d时肺组织p-ERK蛋白分布(Masson染色,×200)
2.2各组肺RAC、W/D、胶原纤维含量比较 第3天,H+N组的RAC较A+C组明显降低(P<0.05)。H+N组、H+C组的胶原纤维含量与A+N组、A+C组比较明显提高(P<0.05),H+N组的胶原纤维含量较H+C组更高(P<0.05)。第7、14天,H+N组、H+C组的RAC、W/D值和胶原纤维含量与A+N组、A+C组比较差异有统计学意义(P<0.05),H+N组、H+C组的RAC降低,W/D值和胶原纤维含量增高;与H+C组比较,H+N组RAC更低,W/D值和胶原纤维含量更高(P<0.05),见表1。
2.3各组肺组织p-ERK蛋白分布 免疫组织化学检测结果表明,p-ERK蛋白阳性反应产物在细胞质表达,为棕黄色颗粒状。与A+N组、A+C组比较,高氧暴露3、7、14 d时H+N组肺组织p-ERK平均光密度值均增加,差异有统计学意义(P<0.05)。而H+C组p-ERK平均光密度值较H+N组明显减少,差异有统计学意义(P<0.05),见图2~3。
表1 各组肺组织RAC、W/D值、胶原纤维含量比较
a:P<0.05,与A+N组比较;b:P<0.05,与A+C组比较;c:P<0.05,与H+N组比较
2.4各组肺组织p-ERK蛋白水平 Western blot结果表明,各组在3个不同的时间段ERK蛋白水平无明显差异(图4B)。高氧暴露第3天、第7天和第14天时,p-ERK蛋白水平在H+N组中明显升高,与A+C组、A+N组比较,差异有统计学意义(P<0.05),其中在第7天水平最高;在高氧暴露第7天和第14天时,H+C组中的p-ERK蛋白水平降低,与H+N组比较差异有统计学意义(P<0.05),见图4C。各组p-ERK/ERK比值与p-ERK蛋白水平情况表现出相同的趋势,见图4D。
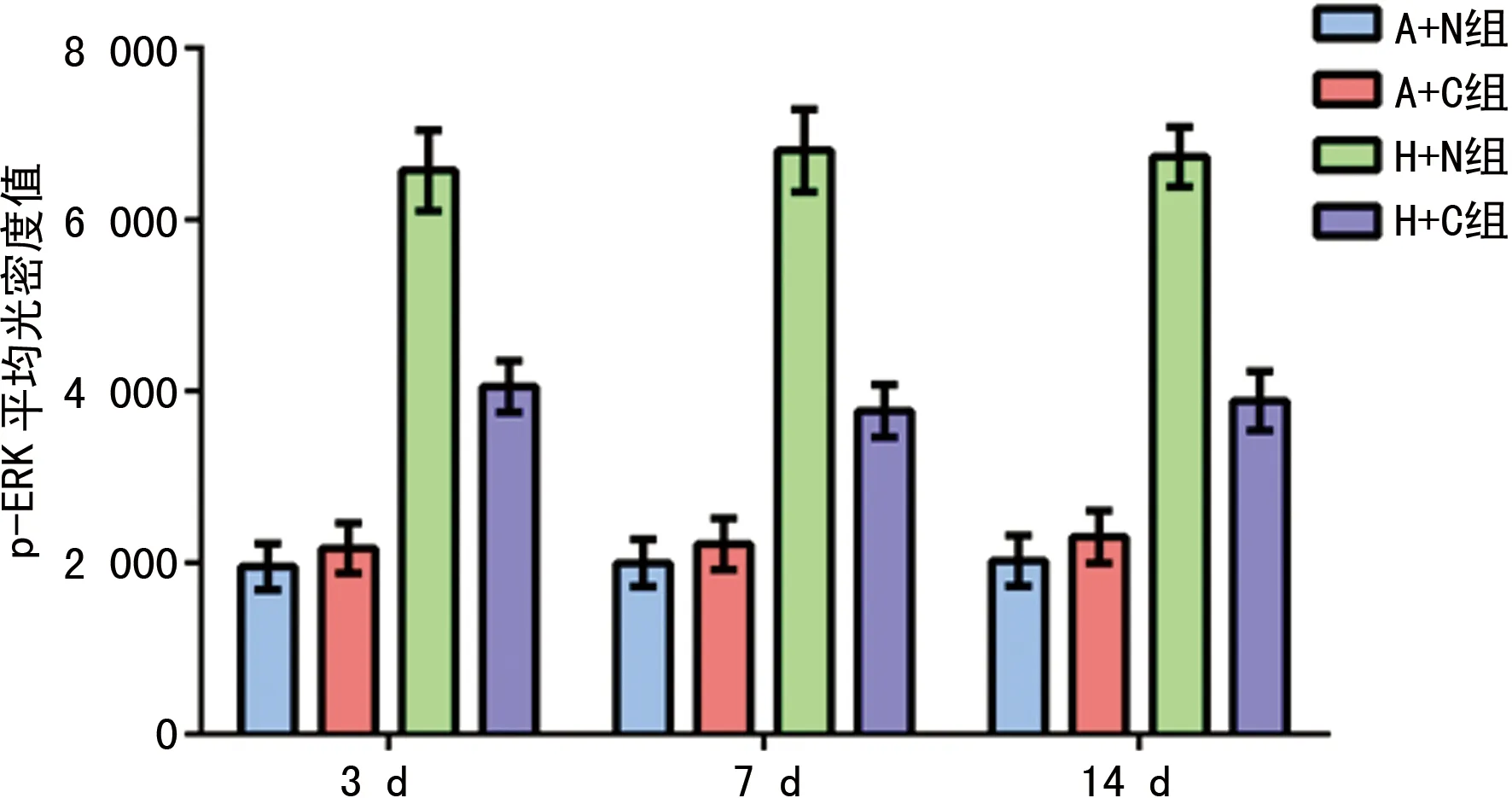
图3 各组3、7、14 d时肺组织p-ERK的平均光密度值

A:ERK、P-ERK、β-actin蛋白灰度条带;B:4组ERK灰度值与β-actin灰度值的比值柱形图;C:4组p-ERK灰度值与β-actin灰度值的比值柱形图;D:4组p-ERK/ERK蛋白相对表达量柱状图
图4 各组ERK和p-ERK蛋白在肺组织中的表达
3 讨 论
随着医疗技术的发展,氧疗及机械通气的应用使早产儿存活率明显提升,但过度或高浓度的氧气治疗极易导致BPD发生。近年来BPD在存活的早产儿中发生率为60%以上,严重影响儿童的生存质量甚至导致其死亡。拮抗氧化应激,抑制肺组织纤维化被认为是减少BPD发生的重要因素,因此本研究拟对咖啡因干预早产鼠高氧肺损伤修复的作用及机制进行研究。本课题组选用妊娠19 d破宫产仔鼠作为实验动物,暴露于高氧环境后,其组织学变化模拟了早产儿的BPD[9]。通过病理切片发现,A+N组与A+C组中,肺组织随着时间延长逐渐发育;高氧暴露7 d时,H+N组肺泡壁增厚,肺组织结构紊乱,肺间质细胞增加,肺泡内大量红细胞及液体渗出,炎性细胞增多,随着高氧暴露时间的延长,H+N组肺间质增厚并伴随纤维化;咖啡因干预后,这一现象得以缓解。这与谭利平等[10]研究相符。
咖啡因是一种非选择性的磷酸二酯酶(PDE)抑制剂,其在治疗预防BPD中的益处是近几年才被学界所发现的[11-12]。JING等[13]研究证实咖啡因通过改变三磷酸鸟苷环化水解酶(GCH1)和四氢生物蝶呤(BH4)水平,进而改善内皮一氧化氮合酶(eNOS)活性来保护新生仔鼠未成熟的肺免于高氧诱导的损伤。在家兔建立的高氧肺损伤模型中,也证实了咖啡因可减少肺部功能性和炎性改变[14]。本研究发现,高氧暴露可导致仔鼠肺组织中RAC降低,W/D值与胶原纤维含量增加,而咖啡因干预后可明显抑制这些现象,这充分说明高氧暴露可增加早产仔鼠肺部炎性反应和肺组织纤维化,使肺发育受阻,咖啡因干预后炎性反应和肺组织纤维化程度减轻。
MAPKs是信号转导途径中重要的中间体,包括ERK、c-Jun、P38和ERK5 4种不同级联反应,ERK作为其重要成员广泛参与细胞生长、增殖、分化、应激反应和细胞凋亡。高氧刺激可以激活ERK,这种现象通常归因于细胞活性氧(ROS)生成[15]。LI等[16]利用吉非替尼抑制ERK激酶磷酸化拮抗博莱霉素诱导的肺纤维化,说明ERK信号通路激活可以使肺组织纤维化。因此,能否抑制ERK信号通路的激活成为拮抗高氧诱导肺损伤的关键因素。本实验通过免疫组织化学和Western blot检测发现,高氧暴露后p-ERK水平明显升高,咖啡因干预使p-ERK水平降低,说明高氧暴露使ERK信号通路激活,而咖啡因可以抑制这一现象。综合病理切片、RAC、W/D值和胶原纤维含量结果,本研究认为高氧暴露后,ERK信号通路被激活,导致肺组织出现炎性反应,肺纤维化程度增加;咖啡因干预后,通过降低p-ERK水平,抑制ERK信号通路激活,从而抑制肺组织纤维化来行使在早产仔鼠高氧肺损伤时的保护作用。
本研究虽然明确了咖啡因可通过抑制ERK信号通路的激活以起到在高氧肺损伤中的保护作用,但并未探讨咖啡因是通过何种途径对p-ERK水平产生影响。在今后的工作中,本课题组将着重探讨咖啡因影响ERK信号通路行使保护作用的具体机制,为临床防治BPD提供更多理论依据。
