旱作栽培条件下源库调控对冬小麦茎秆节间同化物转运的影响
2019-04-16李红兵邓西平
陈 炜,李红兵,邓西平
(1.陕西省灾害监测与机理模拟重点实验室,宝鸡文理学院 地理与环境学院,陕西宝鸡 721013;2.中国科学院 水利部水土保持研究所黄土高原土壤侵蚀与旱地农业国家重点实验室,陕西杨凌 712100)
源库关系协调发展是小麦(TriticumaestivumL.)高产的生物学基础,充足的源和充实的库是小麦高产的必要条件[1]。明确冬小麦籽粒灌浆过程中源端或库端对籽粒产量的影响,对提高冬小麦产量有重要意义[2]。源器官通常是指生产或输出同化物的器官或组织,小麦植株的源系统由绿色的茎、鞘、叶组成。叶片是主要的光合源,是籽粒形成的物质基础[3]。库器官是接受和贮藏同化物的器官或组织,在小麦灌浆过程中,库器官主要指籽粒[4]。籽粒产量的形成是由光合同化物在源端与库端合理分配共同决定的,灌浆过程中源器官制造光合产物,并以蔗糖形式运输到库器官(籽粒),最终在一系列酶的催化下形成淀粉[5]。
人工调控源库关系是研究源库关系的主要方法[6-8],剪叶或遮光是主要的减源方式[9],而疏花是主要的缩库手段[10]。源库关系对冬小麦产量形成的影响与其基因型和生长环境有关[1, 11-12]。目前,主要的缩库手段是将穗部一侧小穗去除,由于不同小穗位于不同的穗位,造成粒质量分布的不均衡[13],处于中部和基部小穗的籽粒,粒质量较高常被定义为强势粒,而处于上部小穗的籽粒,粒质量较轻常被定义为弱势粒[14],即同时去除强势粒和弱势粒。然而,同时去除强、弱势粒会引起籽粒激素含量的变化,影响其同化物的积累[14]。关于去除不同穗位(强势粒或/和弱势粒)籽粒,对源库关系的影响和对茎秆同化物积累转运的研究较少[15]。
用于籽粒灌浆的光合同化物,根据其形成的不同时间可分为花前和花后固定的光合同化物[16-17]。灌浆过程中,光合同化物从源器官向库器官中的转运对籽粒产量的形成有十分重要的作用[18-19]。籽粒产量的形成可被认为是源器官提供光合同化物和库器官积累光合同化物之间相互平衡的结果,源端光合同化物提供不足会造成籽粒质量的降低,而库器官对光合同化物积累能力的不足会引起光合同化物在源器官中的滞留,导致收获指数的降低[20]。水分亏缺是限制旱地冬小麦产量的主要因素,灌浆期干旱引起叶片光合能力的降低,抑制叶片光合作用的进行,增加茎秆中同化物向籽粒的运输,以缓解叶片光合产物供应的不足。而冬小麦非叶器官(穗部和茎秆)对干旱的适应程度高于叶片,是干旱条件下灌浆期同化物的重要来源[21-22]。Ehdaie等[23]将茎秆分为3个不同节间,分析茎秆不同节间对籽粒质量的贡献率。Gebbing等[24]研究表明花前存储在茎秆中的光合同化物对籽粒的贡献率为25%~33%。然而,由于茎秆不同节间在同化物积累[25-26]和光合能力[27]上的差异,为了准确描述旱作冬小麦灌浆过程中,茎秆中同化物积累和转运对不同源库调控的响应。本研究选用旱地冬小麦品种‘长武134’为供试材料,重点探讨不同源库调控对旱地冬小麦灌浆过程中同化物积累和转运的影响,为提高旱地冬小麦对光合同化物的利用提供理论依据。
1 材料与方法
1.1 试验设计
试验于2012年9月至2013年7月在黄土高原中部陕西省长武县洪家镇王东村中国科学院长武农业生态试验站进行,地理位置107°40′E30°14′N,海拔1 200 m,年均降水577 mm,年均气温9.1 ℃,≥10 ℃的积温3 029 ℃,无霜期171 d。该地区冬小麦种植属典型旱作雨养农业体系,土壤类型为黑垆土,质地均匀,疏松多孔,耕性良好。试验田土壤0~20 cm肥力状况为:有机质质量分数11.5 g·kg-1,全氮质量分数0.89 g·kg-1,速效磷质量分数15.4 g·kg-1,速效钾质量分数144.8 g·kg-1。2012-2013年月平均温度和降雨如图1所示。试验当季降雨量共422.5 mm,与多年平均(1957-2006的579.8 mm)相比较,全年降雨量减少27%,而冬小麦花前至灌浆期(5月-6月)的降雨量低于多年平均35%,表明冬小麦在灌浆期受到一定程度干旱胁迫。
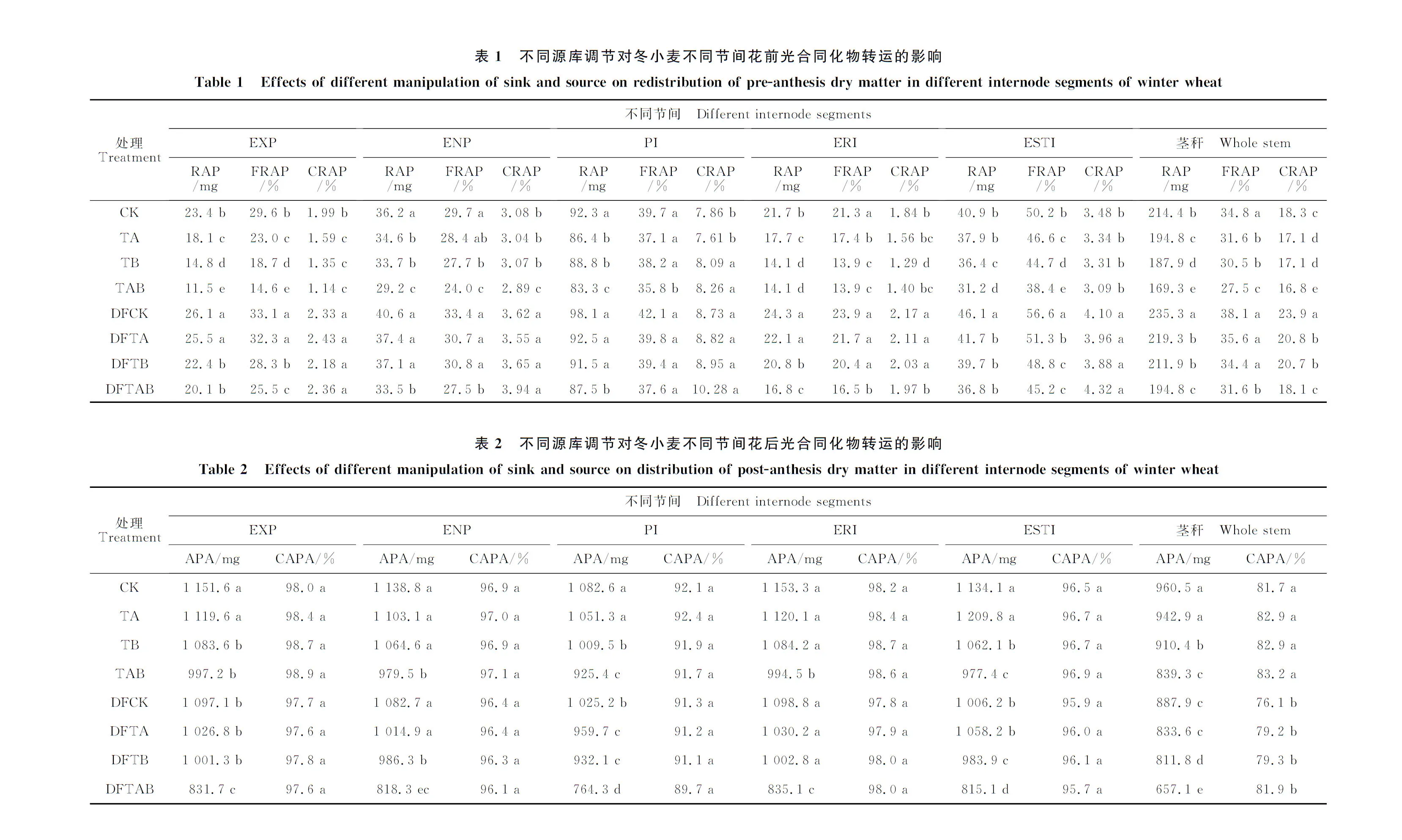
试验以旱地冬小麦品种‘长武134’为供试材料,于2012-2013年进行,小区面积为144 m2(24 m×6 m),小区间隔2 m,4次重复,随机区组设计。施氮量和施磷量分别为150和120 kg·hm-2,其中氮肥以尿素,磷肥以过磷酸钙的形式在播种时作为底肥一次施入。于2012-09-25以人工播种的方式进行种植,行距20 cm,全生育期无灌溉。为揭示不同源库调控对冬小麦茎秆不同节间同化物转运的影响,设置4种缩库处理,分别为去除上4小穗(TA)、去除下4小穗(TB)、同时去除上4小穗和下4小穗(TAB)和完整小穗(CK)。同时设置剪叶减源处理(去除旗叶,DF),并在DF处理下设置缩库处理,分别记作DFCK(去除旗叶+完整小穗),DFTA(去除旗叶+上4小穗),DFTB(去除旗叶+下4小穗)和DFTAB(去除旗叶+上4小穗和下4小穗)共8种源库调控方式,在上述缩库处理中,穗轴均被保留。同时按照图2的方式,将冬小麦茎秆分为5个不同的节间,穗下节间可见光部分(Exposed peduncle of internode,EXP),穗下节间不见光部分(Enclosed by flag leaf of peduncle,ENP),倒二节间(Penultimate internode,PI),倒三节间(Lower internode,ERI),倒四节间(Lowest internode,ESTI)。

图1 试验当季(2012-2013)与多年平均(1957-2006)降水量和温度Fig.1 Precipitation and temperature in 2012-2013 and the long-term (1957-2006) means
1.2 采样与指标测定
1.2.1 采样 每小区选择50个生长一致的单茎进行标记,并在开花当天进行疏花缩库和剪叶减源处理,分别于开花当天和收获期取样。将冬小麦分为穗部和茎秆,其中将穗部剥离成穗(成熟期分离出籽粒),将茎秆按照图2的方式分为不同的节间,分别装入纸袋,然后放入105 ℃烘箱中杀青30 min,再降至60 ℃烘至恒量,称量。其中对单穗质量进行称量时均不包括穗轴。
1.2.2 不同节间同化物转运对籽粒产量贡献率 按照Jiang等[28]的方法分别计算茎秆不同节间、花前和花后同化物对冬小麦单穗质量的贡献:不同节间花前同化物的转运量(RAP)、转运效率(FRAP);不同节间花前同化物对籽粒的贡献率(CRAP)、不同节间花后同化物的转运量(APA)和不同节间花后同化物对籽粒的贡献率(CAPA)。RAP=特定节间开花期干质量-特定节间成熟期干质量;FRAP=(特定节间开花期干质量-特定节间成熟期的干质量)/特定节间开花期干质量×100%;CRAP=(特定节间开花期干质量 - 特定节间成熟期的干质量)/成熟期籽粒质量×100%;APA=成熟期籽粒质量 - (特定节间开花期干质量 - 特定节间成熟期干质量)/成熟期籽粒质量;CAPA=特定节间花后同化物转运量/成熟期籽粒质量×100%。
1.3 数据处理
采用Microsoft Excel 2010对数据进行处理,用Sigmaplot 12.5绘图;用SPSS 20.0进行差异显著性检验。
2 结果与分析
2.1 不同源库调节对冬小麦单穗质量的影响
不同源库调控处理显著影响冬小麦单穗质量(图3)。收获期单穗质量为0.85~1.17 g。减源处理(DF)显著降低单穗质量,与相应的对照(完整旗叶比较),单穗质量的平均减幅为9.1%,其中以TAB和DFTAB间的差异最大,为18%,而TB和DFTB间的差异最小,为7%。可能是由于花后去除旗叶(主要的叶光合源),降低了光合同化物的积累,最终单穗质量低于相应完整旗叶中的单穗质量。不同缩库处理之间,随着缩库强度的增加,其单穗质量逐渐降低,呈现出CK>TA>TB>TAB的变化趋势。CK单穗质量比TAB单穗质量高16%;而DFCK和DFTAB单穗质量之间的差异为31.9%。

不同大写字母表示不去旗叶处理间差异显著(P<0.05) Different capital letters in the same column indicate significant difference among whole flay leaf treatment(P<0.05);小写字母表示减源处理间差异显著(P<0.05) Different lowercase letters in the same column indicate significant difference among defoliate flag leaf treatments(P<0.05);*表示完整旗叶和减源处理之间差异显著(P<0.05) * indicate significant difference between whole flag leaf treatment and reduce sinx treatment;下同 the same below
图3不同源库调节对冬小麦单穗质量的影响
Fig.3Effectsofdifferentsinkandsourcemanipulationonthesinglespikemassofwinterwheat
2.2 不同源库调节对冬小麦平均粒质量的影响
由图4可知,减源处理(DF),降低平均粒质量。与对应CK相比较,DF平均降低粒质量为6%,其中DFTAB和TAB间的减幅最大为8%,而DFCK和CK间的减幅最小为3%,花后去除旗叶,影响花后光合作用的进行,降低了光合同化物的积累,显著降低了平均粒质量。不同缩库处理对冬小麦平均粒质量的影响呈现出TAB>TA>CK>TB的趋势。其中,TAB由于同时去除了上4小穗和下4小穗(过度缩库),使灌浆过程中更多的光合同化物分布到剩余的籽粒中,同时中部小穗主要由强势粒所构成,增加了中部强势粒的填充,有助于平均粒质量的增加。虽然TA与TB均为去除4个小穗,但由于去除的小穗位置不同,引起剩余籽粒对光合同化物在利用上的差异。其中,TA去除的为上4小穗,而上4小穗主要由弱势粒构成,用于填充上4小穗籽粒的光合同化物较少,引起较多的光合同化物填充中部和下部小穗,同时中部和下部小穗主要由强势粒构成,而作为强势粒,其库端对籽粒质量提升的限制较小,最终有助于其粒质量的增加;而TB去除的为下4小穗,下4小穗主要由强势粒构成,用于填充该部位籽粒所需的光合同化物较多,使较少的光合同化物用于填充上部和中部小穗,同时上部小穗主要由弱势粒构成,其库端对籽粒粒质量的提高有一定的限制,因此TB处理平均粒质量低于TA和CK。此外在TA中,去除上部小穗籽粒的同时,可能降低了弱势粒中激素对剩余籽粒质量增加的限制,以及花后穗部仍可进行光合作用,而光合作用与光合面积有关,而上4小穗面积小于下4小穗面积,在一定程度上TA处理下,穗部籽粒光合积累的同化物大于TB处理。因此导致TA处理平均粒质量大于CK和TB处理。
2.3 不同源库调控对冬小麦茎秆不同节间花前光合同化物转运的影响
不同源库调控显著影响茎秆不同节间花前光合同化物的积累、转运和对籽粒的贡献率(表1)。在所有处理中,DFCK花前贡献率最高为23.9%,而TAB花前贡献率最低为16.8%。与CK比较,DF处理增加花前茎秆光合同化物的转运量、转运效率和贡献率,增幅分别为12%、12%和20%。表明由于去除主要的光合叶器官(旗叶),冬小麦为了维持籽粒质量的积累,动员较多的花前光合同化物以弥补因去除旗叶所引起的光合同化物供应的不足,即冬小麦产生补偿效应,以维持籽粒的质量。不同缩库处理之间,随着缩库程度的增加,RAP、FRAP和CRAP都逐渐降低,表现出CK>TA>TB>TAB的趋势,可能是由于随着缩库强度的增加,导致籽粒对光合同化物需求减少,引起较多的光合同化物滞留在茎秆中,最终引起花前光合同化物转运的降低。茎秆不同节间中,以倒二节间的花前贡献率最大,可能是由于倒二节间具有较大的干质量,而穗下节间可见光部分花前贡献率最低,可能是由于穗下节间可见光部分质量较低,同时,该节间处于茎秆的最顶部,为了降低倒伏的概率,因此其质量最低,最终导致花前贡献率最低。虽然下部节间(倒三节间和倒四节间)FRAP较高,但其CRAP却不是最高,可能是由于下部节间将一部分的光合同化物转运到其他节间中,用于节间的生长,而没有直接运输到籽粒中,形成籽粒质量。

图4 不同源库调节对冬小麦平均粒质量的影响Fig.4 Effects of different sink and source manipulation on the average grain mass of winter wheat
2.4 不同源库调控对冬小麦茎秆不同节间花后光合同化物转运的影响
不同源库调控对茎秆不同节间花后光合同化物的影响如表2所示。不同源库调控中,以DFCK处理CAPA最低为76.1%,而TAB处理CAPA最高为83.2%。随着旗叶的去除,降低了光合作用的进行,导致花后光合作用所固定的同化物降低,最终引起花后光合同化物对籽粒的贡献率降低。随着缩库程度的增加,其花后光合同化物对籽粒的贡献率逐渐增加,但处理间差异不显著。从可见光到倒二节间花后光合同化物对籽粒的贡献率逐渐降低,可能由于茎秆作为非叶光合器官,不同节间从上至下接受光照的概率和CO2浓度逐渐降低,最终影响花后茎秆光合作用的进行。
3 讨 论
花后小麦光合能力(源)和籽粒对光合产物转化贮藏能力(库)成为制约小麦产量的两个重要因素,而减源缩库是研究小麦源库关系最常用的一种手段。本试验结果显示,减源处理(DF)降低了冬小麦单穗质量和平均粒质量。主要是因为随着旗叶的去除,降低了花后光合作用,减少了光合同化物的积累,导致花后光合同化物对籽粒贡献率的降低。茎秆作为光合同化物主要的存储器官对维持籽粒的产量具有十分重要的作用[29]。灌浆期茎秆中存储的光合同化物可分为两类,一类是开花前积累并存储在茎秆中的光合同化物,另一类是花后光合作用所固定的光合同化物。前者对维持籽粒灌浆有重要的作用,后者可以储存在源器官或者直接转运到库中[22, 30]。本试验减源处理中,虽然花后光合同化物对籽粒的贡献率低于对照,但是却增加了茎秆中花前光合同化物的转运效率和对籽粒的贡献率,表明通过增加花前光合同化物的运转,在一定程度上能够弥补由于减源处理导致的单穗质量和平均粒质量的降低,但是茎秆花前光合同化物的转运并不能完全弥补叶源亏缺造成的质量损失。说明,花后光合同化物对籽粒产量的形成起主要作用[16]。
本试验中,不同缩库处理之间,单穗质量呈现出CK>TA>TB>TAB的变化趋势,随着缩库程度的加剧,单穗质量逐渐降低。然而,平均粒质量呈现出逐渐增加的趋势。可能是随着缩库程度的加剧,使剩余籽粒获得光合同化物的量发生了改变。单穗质量是由粒质量和穗粒数两者共同决定[31],虽然不同程度的缩库均增加了平均粒质量,但是其单穗质量却低于完整小穗,表明由于穗粒数的降低引起单穗质量的损失,没有通过增加平均粒质量得到弥补。过度的缩库处理(TAB),降低了穗粒数,缓解了籽粒对同化物的竞争,最终引起平均粒质量的增加。虽然TA和TB均是去除4个小穗,但上述2种处理,对剩余籽粒质量的影响呈现显著的穗位性。分析TA和TB发现,前者去除的是上4小穗,使较多的光合同化物分配到剩余籽粒中,引起剩余籽粒对同化物竞争性利用的强度小于后者(去除下4小穗),最终导致TA中部小穗粒质量大于TB中部小穗籽质量。同时TA去除的为上4小穗,而上4小穗主要由弱势粒所构成[32],弱势粒中较高的激素浓度(主要是脱落酸),显著降低其粒质量的增加[33],而去除上4小穗之后,有可能同时去除了高浓度激素对籽粒质量增加的限制,进一步引起TA处理中剩余籽粒质量的增加。同时还可能与剪穗程度有关,因为穗自身的光合作用对于籽粒质量的累积也有重要的作用。
茎秆不同节间不同类型的光合同化物对籽粒质量的贡献呈现出不同的特点,下部节间花前光合同化物对籽粒的贡献率大于上部节间花前光合同化物对籽粒的贡献率,这个结果与已有的研究结果相一致[34],虽然倒三节间和倒四节间其花前光合同化物转运效率较高,但对籽粒产量的贡献率较低,可能是由于上述2节间将一部分光合同化物转运到其他节间,而没有直接转运到籽粒中。茎秆不同节间,花后光合同化物对籽粒的贡献率呈现出与花前光合同化物对籽粒的贡献率相反的趋势,从可见光部分到倒二节间,花后光合同化物对籽粒的贡献率逐渐降低,可见光部分具有最高的花后光合同化物贡献率,其次为不见光部分,然后为倒二节间。可能是由于茎秆自上而下,光照强度和CO2浓度逐渐降低,同时可见光部分属自养器官,且有较高的磷酸烯醇式丙酮酸羧化酶的活力[26],有助于其花后光合同化物对籽粒贡献率的提高。
在黄土高原长武地区,有研究表明随着冬小麦品种的演替,冬小麦叶片光合作用对籽粒产量的贡献较低[35-36],要获得冬小麦高产,源库协调发展是重要影响因素,虽然花前光合同化物转运对籽粒产量的形成有一定的贡献,但花后光合作用对籽粒产量的形成起主要作用,而该地区冬小麦灌浆过程往往伴随着干旱胁迫,因此在该地区冬小麦的栽培过程中应注重源器官在光合同化物积累上的增强。在旱作条件下,旗叶易受到水分胁迫的影响,降低光合同化物的积累能力,而茎秆和穗部作为主要的非叶光合器官,对维持光合作用的进行有重要作用,有研究表明,穗部的光合作用对维持粒质量的增加有十分重要的作用,特别是在水分亏缺的情况下[37]。通过增加非叶器官的光合作用,可以提高灌浆过程中光合同化物的积累,其中穗部不仅在决定穗粒数上起到重要的作用,同时又是主要的非叶光合器官,因此在长武地区穗部可以作为提升籽粒产量的潜在器官。对于冬小麦不同节间同化物转运对籽粒产量的贡献率可通过同位素标记的方法进一步研究,以便更好地分析不同节间在同化物积累转运的特点。
