车桑子小孢子发生与雄配子体发育
2019-04-04卢雪佳刘方炎高成杰李昆
卢雪佳, 刘方炎, 高成杰, 李昆
车桑子小孢子发生与雄配子体发育
卢雪佳, 刘方炎, 高成杰, 李昆*
(中国林业科学研究院资源昆虫研究所,昆明 650224)
为了解干热河谷区车桑子()胚胎学特征及其结籽率低的原因,采用常规石蜡切片法和电镜扫描技术对车桑子小孢子发生、雄配子体发育和花粉的形态特征进行了观察。结果表明,车桑子花药具有4个花粉囊。完整的花药壁从外到内依次为表皮、药室内壁、2~3层中层细胞和绒毡层; 绒毡层类型是腺质绒毡层。花药成熟期,中层、绒毡层均退化消失。小孢子母细胞进行同时型胞质分裂;四分体为四面体型结构。成熟的花粉为二细胞型。花粉近球形,外壁密布颗粒状纹饰,具有3条不构成合沟的萌发沟。雄性生殖发育过程出现的异常现象可能是干热河谷地区车桑子结籽率低的原因之一。
车桑子;小孢子;雄配子体;花粉;形态特征
车桑子()是无患子科(Sapin- daceae)车桑子属的灌木或小乔木,广泛分布于世界的热带和亚热带地区,我国主要分布于西南部、南部至东南部。以其为优势种组成的稀树灌丛、灌丛或灌草丛是干热河谷地区几种重要的优势乡土植被[1]。车桑子具有耐干旱、萌生能力强、根系发达和丛生性等生长习性,常生长于沙土、干旱山坡、旷地等处,是一种良好的保水固土植物[2],常被用作我国西南干热河谷地区造林的先锋树种,在困难立地植被恢复方面发挥着重要作用,具有较高的生态价值。
目前,对车桑子的研究主要集中在造林树种的筛选[3]、光合特性及光合水分生理特性[4]、干旱胁迫下种子和幼苗生理指标的变化[5]、种子的萌发特性和影响幼苗生长发育的生态因子[6–7]、旱生结构特征[8]等。然而,有关车桑子繁殖特征和繁殖生物学的研究报道较少。本试验采用常规石蜡切片和电镜扫描技术对车桑子的小孢子发生和雄配子体的发育,以及花粉粒的形态特征进行了观察,为车桑子的生殖生物学和发育生物学研究提供基础资料,并试图从生殖生物学方面分析其结籽率低的原因,为车桑子的开发利用提供理论依据。
1 材料和方法
1.1 材料
车桑子()采自云南省楚雄自治州元谋县国家林业局元谋荒漠生态系统定位研究站附近干热河谷区域。选取植株中不同发育阶段的雄花花蕾和花朵,保存于FAA固定液(50%酒精90 mL∶甲醛5 mL∶冰醋酸5 mL)中,4℃冰箱中保存备用。凭证标本可见于中国科学院昆明植物研究所标本馆(KUN)的蜡叶标本,采集人为滇西金沙江队,采集号为6507。
1.2 方法
石蜡切片 从固定液中取出花蕾,经系列脱水、二甲苯透明、浸蜡、包埋。用切片机进行连续切片,切片厚度为6~8m,番红-固绿进行对染, 蔡司光学显微镜下观察并拍照。
电镜扫描 取即将开放的花蕾,剥离出成熟花药,轻轻划破花药表皮,挤压出花粉粘于双面导电胶布上,置于金属样品台进行喷金,在扫描电镜下观察并进行拍照。
2 结果和分析
2.1 花药壁的发育
车桑子的花药具有4个花粉囊,分为左、右两半,中间由药隔相连,花药中4个花粉囊孢原细胞的出现具有不同步性(图1: A)。在花药4个角隅处表皮下均有较大体积、细胞核明显的孢原细胞(图1: B)。然后,孢原细胞经过1次平周分裂,形成两层细胞,即外层的初生壁细胞和内层的造孢细胞(图1: C)。初生壁细胞继续进行数次平周分裂形成4~5层细胞,最外层紧贴表皮的一层细胞为药室内壁, 中间为2~3层中层,最内层为绒毡层。由此可见, 完整的花药壁由外向内依次为表皮细胞(1层)、药室内壁细胞(1层)、中层细胞(2~3层)和绒毡层细胞(1层), 共5~6层细胞组成(图1: D, E)。根据Davis的分类[9], 车桑子的花药壁发育属于基本型。
在花药逐渐成熟时,中层细胞受到挤压变形, 绒毡层细胞出现退化解体的现象(图1: J)。根据胡适宜的划分,车桑子的绒毡层细胞是原位解体,属于腺质绒毡层[10]。当花药成熟时,同侧两个花粉囊之间的花粉囊壁破裂,两个花粉囊相互沟通(图1: K)。成熟的车桑子花药开裂方式为纵裂,裂口位于两花粉囊交界处(图1: L)。散粉时,花粉囊壁仅剩下表皮细胞残余物和一层加厚伸长的纤维层(图1: M)。
2.2 小孢子发生
孢原细胞进行平周分裂产生的内层造孢细胞进行多次分裂,形成次生造孢细胞。次生造孢细胞的细胞核较大、核仁明显,细胞形状呈多边形,排列非常紧密(图1: D)。次生造孢细胞发育形成小孢子母细胞,此时小孢子母细胞体积变大,逐渐趋于圆形,细胞质较浓、细胞核较大、核仁明显,为进入减数分裂期做准备,此时绒毡层细胞正常(图1: E)。当小孢子母细胞连续进行两次细胞分裂后,形成了4个排列紧密的子细胞且被包围在胼胝质内,即四分体(图1: F),4个子细胞的排列方式为四面体,同时中层细胞受到挤压逐渐退化解体,绒毡层细胞开始部分出现解体现象。小孢子母细胞的整个减数分裂过程均被包围在胼胝质壁中。
2.3 雄配子体的发育
四分体后期,在胼胝质酶的作用下,胼胝质壁逐渐被溶解,四分体中的细胞彼此分离,形成了4个单核的花粉粒细胞。刚分离的单核花粉粒细胞的细胞核居中且明显,此时绒毡层解体(图1: G)。随着单核花粉粒细胞逐渐扩大,液泡的挤压,细胞核开始逐渐靠边(图1: H)。此时,花粉粒细胞进行1次不均等的分裂, 形成1个较大的营养细胞和1个较小的生殖细胞(图1: I)。二核花粉粒完全成熟时,绒毡层和中层细胞全部解体。成熟花粉粒上可见萌发沟(图1: I)。扫描电镜下车桑子的花粉粒为近球形, 一般有3条萌发沟,极少见4和2条萌发沟。车桑子花粉粒的3条萌发沟不构成合沟,花粉外壁密布颗粒状的纹饰(图1: P, Q)。极轴直径为(24.91±2.03)m, 赤道轴直径为(23.70±1.64)m。在车桑子雄配子体的发育过程中,观察到部分败育花粉,主要表现出形状各异、花粉粒干瘪、皱缩、无核内物质,但这种花粉只少量出现。另外也观察到部分花粉囊内为空室(图1: N, O)。
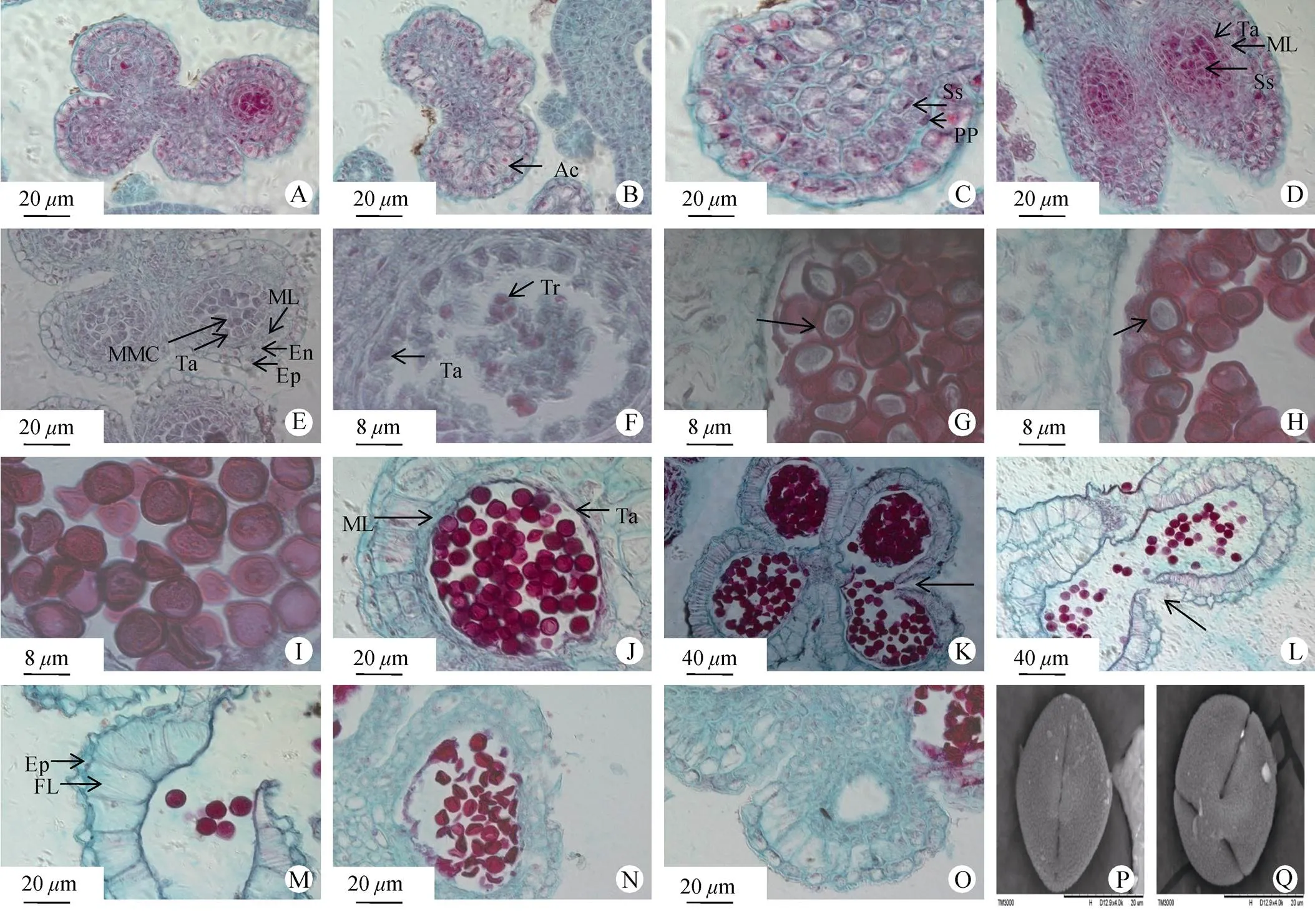
图1 花药发育过程。A: 花药含有4个花粉囊; B: 孢原细胞(箭头); C: 初生壁细胞和造孢细胞(箭头); D: 初生造孢时期; E: 小孢子母细胞时期; F: 四分体; G: 小孢子单核居中期; H: 小孢子单核靠边期; I: 成熟的花粉粒; J: 退化的绒毡层; K: 同侧2个花粉囊相互连通; L: 花药成熟时纵裂,裂口在2个药室的连接处; M: 表皮和纤维层; M: 败育花粉粒; O: 空的花粉囊; P: 成熟花粉粒(极面); Q: 成熟花粉粒(赤道面)。Ac: 孢原细胞; PP: 初生壁细胞; Ss:造孢细胞; Ta: 绒毡层; ML: 中层; MMC: 小孢子母细胞; En: 药室内壁; Ep: 表皮细胞; Tr: 四分体; FL: 纤维层。
3 讨论
本试验观察了车桑子的小孢子发生、雄配子体发育过程和花粉形态,车桑子具有4个花粉囊,横切面呈蝶形,花药壁由5~6层细胞构成,从外至内依次为1层表皮、1层药室内壁、2~3层中层和1层绒毡层,为腺性绒毡层,花药壁的发育类型为基本型。小孢子四分体为四面体型。成熟花粉粒为二核型。部分胚胎学特征与同科植物类似,文冠果()、倒地铃()和茶条木()的小孢子四分体均为四面体型、成熟花粉粒为两核型[11–13]。车桑子花粉粒近球形,多具有3条萌发沟,稀2或4条萌发沟,表面密布颗粒状纹饰,极轴为(24.91± 2.03)m,赤道轴为(23.70±1.64)m。而夏丽敏等[14]的研究表明,广州龙洞地区的车桑子花粉极轴为(26.5±2.1)m、赤道轴为(25.0±1.86)m,本研究中车桑子采于元谋干热河谷地区,其极轴和赤道轴均较小,表明车桑子的花粉大小可能存在区域差异, 且干热河谷区花粉相对较小。本研究所获得的车桑子的胚胎学特征,充实了无患子科胚胎学研究资料。此外,车桑子同一花序中雄蕊发育程度不同步,同一朵雄花中不同花药发育程度不同,同一花药的不同花粉囊发育程度不同步。这种现象在‘早籽银桂’(‘Zao Zi Yin’)、‘晚银桂’(‘Wan Yin’)中也有出现[15–16],可能是为了延长散粉时间,进而增加授粉机会的一种适应性机制。
在花药的发育过程中,绒毡层位于花药壁最内层,直接与小孢子发育相关。绒毡层具有分泌功能,一方面能提供小孢子发育所需营养物质;另一方面能释放胼胝质酶溶解胼胝质壁[17]。绒毡层在花粉发育过程中具有重要作用,绒毡层细胞提前或延迟降解或出现任何异常都有可能导致雄性不育及花粉败育[18–19]。在‘晚银桂’的花药发育过程中出现了绒毡层提前退化现象,导致了花粉败育[16];鹅掌楸()也发生绒毡层细胞异常现象而导致的雄性不育[20]。同科植物倒地铃的绒毡层异常可能导致雌花雄性不育[13],吕雪芹等[21]认为文冠果绒毡层延迟解体或退化不彻底导致可孕花的花药败育。本试验观察到部分绒毡层细胞在四分体时期已经解体,由此推测,车桑子花药发育过程中出现的败育现象,可能与绒毡层提前退化有关。通常,特殊的外界环境条件和遗传特性的原因会引起绒毡层以及雄配子体的异常发育,从而导致花粉败育现象。环境因素导致了连翘()雄配子体不能正常发育进而引起花粉败育[22];毛白杨()的花粉败育则是异质性遗传与环境互作的结果[23]。本研究中,车桑子生长于干湿季分明的干热河谷地区,气候特殊,旱季高温干旱且漫长(当年11月至次年5月),雨季易出现间歇性干旱[24],因此,我们推测车桑子花药壁的绒毡层和雄配子体的异常发育可能与恶劣的环境因素有关,这还有待于进一步研究。
植物的有性生殖是一个复杂的过程,且受内因和外因的共同影响,植物生活史周期的任何一个环节都影响着生殖成功与否[25]。根据调查,车桑子的结籽率较低,仅为(22.5±2.5)%。初步判断,花药发育过程中出现的异常现象可能是导致车桑子结籽率低的原因之一。为进一步确定干热河谷地区车桑子结籽率低、天然更新能力差[24]的原因,本研究将继续从其他生殖生态方面,如花粉量和花粉萌发率、大孢子发生与雌配子体发育是否存在异常、受精过程是否有效、胚的发育是否正常、种子活力高低、种子萌发的状况、环境对种子萌发及幼苗生长的影响等进一步研究。这将为干热河谷区车桑子的进一步开发利用提供一定的理论依据。
致谢 国家林业局元谋荒漠生态系统研究站刘昆友同志帮助采集了部分试验材料,资源昆虫研究所郭晓娟同学及西南林业大学严冬同学在试验过程提供了帮助。
[1] JIN Z Z, OU X K. Vegetation in Yuanjiang, Nujiang, Jinshajiang, Lancangjiang Dry-hot Valley [M]. Kunming: Yunnan University Press, Yunnan Science and Technology Press, 2000: 141–180.金振洲, 欧晓昆. 元江、怒江、金沙江、澜沧江干热河谷植被 [M]. 昆明: 云南大学出版社, 云南科技出版社, 2000: 141–180.
[2] Wang X M, Yan B G, Liu G C, et al. Spatial variation in seed dormancy and germination ofin Yuanmou Dry-hot Valley, China [J]. J Trop Subtrop Bot, 2016, 24(4): 375–380. doi: 10. 11926/j.issn.1005-3395.2016.04.003.王雪梅, 闫帮国, 刘刚才, 等. 元谋干热河谷车桑子种子休眠与萌芽的空间变异特征研究 [J]. 热带亚热带植物学报, 2016, 24(4): 375–380. doi: 10.11926/j.issn.1005-3395.2016.04.003.
[3] LI K, ZHANG C H, CUI Y Z, et al. A study on the fitting afforestation tree species during coverting the land for forestry in hot and arid valley of Jinsha-river [J]. For Res, 2004, 17(5): 555–563. doi: 10.3321/j.issn: 1001-1498.2004.05.002.李昆, 张春华, 崔永忠, 等. 金沙江干热河谷区退耕还林适宜造林树种筛选研究 [J]. 林业科学研究, 2004, 17(5): 555–563. doi: 10. 3321/j.issn:1001-1498.2004.05.002.
[4] LI J Y, JIA L Q, LANG N J, et al. Characteristics of photosynthesis ofin the dry-hot valley of Jinsha River [J]. J Beijing For Univ, 2004, 25(5): 20–24. doi: 10.3321/j.issn:1000-1522.2003.05. 005.李吉跃, 贾利强, 郎南军, 等. 金沙江干热河谷坡柳的光合特性 [J]. 北京林业大学学报, 2003, 25(5): 20–24. doi: 10.3321/j.issn:1000- 1522.2003.05.005.
[5] CHEN S Y, LANG N J, LI J Y, et al. Changes of cytomembrane permeability and proline content in leaves of,andunder drought stress [J]. Guihaia, 2006, 26(1): 80–84. doi: 10.3969/j.issn.1000-3142.2006.01. 019.陈少瑜, 郎南军, 李吉跃, 等. 干旱胁迫下坡柳等幼苗质膜相对透性和脯氨酸含量的变化 [J]. 广西植物, 2006, 26(1): 80–84. doi: 10. 3969/j.issn.1000-3142.2006.01.019.
[6] ZHANG Q Y, SUN H L, LI S C, et al. Different environmental conditions on germination characteristics of(L.) Jacq [J]. Seed, 2013, 32(1): 12–14,19. doi: 10.3969/j.issn.1001-4705. 2013.01.004.张琼瑛, 孙海龙, 李绍才, 等. 不同环境条件对车桑子萌发的影响 [J]. 种子, 2013, 32(1): 12–14,19. doi: 10.3969/j.issn.1001-4705.2013. 01.004.
[7] Wang C H, Zeng C H, Shen D S, et al. Climatic effects of the artificial reservoir of China’s dry-hot valleys on the growth and physio- logical traits ofseedlings [J]. Acta Ecol Sin, 2016, 36(17): 5343–5352. doi: 10.5846/stxb201502110335.王传华, 曾春函, 沈德嵩, 等. 云南干热河谷水库气候效应对车桑子幼苗生长发育的影响及其作用机制 [J]. 生态学报, 2016, 36(17): 5343–5352. doi: 10.5846/stxb201502110335.
[8] GE L, HUANG D, MA H C, et al. Comparison of leaves anatomical structures and related drought resistance among different populations ofin Yunnan hot-dry valley [J]. J Arid Land Resour Environ, 2015, 29(5): 138–143. doi: 10.13448/j.cnki.jalre.2015.166.葛娈, 黄冬, 马焕成, 等. 云南干热河谷不同居群坡柳叶片的旱生结构比较 [J]. 干旱区资源与环境, 2015, 29(5): 138–143. doi: 10. 13448/j.cnki.jalre.2015.166.
[9] Davis G L. Systematic Embryology of the Angiosperms [M]. New York: John Wiley & Sons, 1966: 6–17.
[10] HU S Y. Reproductive Biology of Angiosperms [M]. Beijing: Higher Education Press, 2005: 29–69.胡适宜. 被子植物生殖生物学 [M]. 北京: 高等教育出版社, 2005: 29–69.
[11] CAO L M, CAO M, LIU J H, et al. Sporogenesis and gametogenesis of(Sapindaceae) and their systematic implications [J]. J Syst Evol, 2014, 52(4): 533–539. doi: 10.1111/jse.12083.
[12] FENG X K. A preliminary study on the formation of megaspores, microspores, embryos and endosperm ofBunge [J]. For Sci Technol, 1984(2): 15–18. doi: 10.13456/j.cnki.lykt.1984. 02.006.冯显逵. 文冠果大、小孢子及胚和胚乳形成过程的初步研究 [J]. 林业科技通讯, 1984(2): 15–18. doi: 10.13456/j.cnki.lykt.1984.02.006.
[13] TONG P P, WANG L L, LIU W Z. Study on the morphodifferentiation and anther structure in flowers ofLinn [J]. Plant Sci J, 2016, 34(2): 165–174. doi: 10.11913/PSJ.2095-0837. 2016.20165.仝盼盼, 王玲丽, 刘文哲. 倒地铃花的形态分化与花药结构研究[J].植物科学学报, 2016, 34(2): 165–174. doi: 10.11913/PSJ.2095-0837. 2016.20165.
[14] CAO L M, XIA N H, CAO M, et al. Pollen morphology and its syste- matic significance in Sapindaceae from China [J]. Plant Sci J, 2016, 34(6): 821–833. doi: 10.11913/PSJ.2095-0837.2016.60821.曹丽敏, 夏念和, 曹明, 等. 中国无患子科的花粉形态及其系统学意义 [J]. 植物科学学报, 2016, 34(6): 821–833. doi: 10.11913/PSJ. 2095-0837.2016.60821.
[15] DONG L G, DING Y L. Development of micro- and mega-spores and morphology of floral organ in‘Zao Zi Yin’ [J]. J Trop Subtrop Bot, 2017, 25(3): 299–308. doi: 10.11926/jtsb.3655.董立格, 丁雨龙. ‘早籽银桂’大小孢子发育与花器形态变化 [J]. 热带亚热带植物学报, 2017, 25(3): 299–308. doi: 10.11926/jtsb.3655.
[16] DONG L G, DING Y L, WANG X R, et al. The microsporogenesis and the development of male gametophyte in‘Wanyin- gui’ [J]. J Nanjing For Univ (Nat Sci), 2015, 39(4): 44–50. doi: 10. 3969/j.issn.1000-2006.2015.04.008.董立格, 丁雨龙, 王贤荣, 等. ‘晚银桂’的小孢子发生和雄配子体发育 [J]. 南京林业大学学报(自然科学版), 2015, 39(4): 44–50. doi: 10. 3969/j.issn.1000-2006.2015.04.008.
[17] LI Y X, LI L, CHEN G H. Advances in tapetum development of anthers in higher plants [J]. Crop Res, 2009, 23(5): 287–289. doi: 10. 3969/j.issn.1001-5280.2009.z1.001.李懿星, 李莉, 陈光辉. 高等植物花药绒毡层发育研究进展 [J]. 作物研究, 2009, 23(5): 287–289. doi: 10.3969/j.issn.1001-5280.2009.z1. 001.
[18] ZHANG H, LIANG W Q, ZHANG D B. Research progress on tapetum programmed cell death [J]. J Shanghai Jiaotong Univ (Agric Sci), 2008, 26(1): 86–90. doi: 10.3969/j.issn.1671-9964.2008.01.021.张虹, 梁婉琪, 张大兵. 花药绒毡层细胞程序性死亡研究进展 [J]. 上海交通大学学报(农业科学版), 2008, 26(1): 86–90. doi: 10.3969/ j.issn.1671-9964.2008.01.021.
[19] LUO H S, MENG D X, CHEN X Y, et al. Advances in mechanism of pollen abortion caused by abnormal tapetum in plant [J]. Crop Res, 2012, 26(6): 725–729. doi: 10.3969/j.issn.1001-5280.2012.06.28.罗海山, 孟德璇, 陈晓阳, 等. 植物绒毡层异常导致花粉败育的机理研究进展 [J]. 作物研究, 2012, 26(6): 725–729. doi: 10.3969/j.issn. 1001-5280.2012.06.28.
[20] YIN Z F, FAN R W. The ultrastructure observation on the abortion ofpollen [J]. J Plant Resour Environ, 1997, 6(2): 1–7.尹增芳, 樊汝汶. 鹅掌楸花粉败育过程的超微结构观察 [J]. 植物资源与环境, 1997, 6(2): 1–7.
[21] LÜ X Q, ZHANG M, WANG D, et al. Comparative study on the bisexual flower and unisexual male flower ofBunge [J]. Bull Bot Res, 2014, 34(1): 85–94. doi: 10.7525/j.issn.1673- 5102.2014.01.012.吕雪芹, 张敏, 王頔, 等. 文冠果可孕花与不孕花发育过程的比较研究 [J]. 植物研究, 2014, 34(1): 85–94. doi: 10.7525/j.issn.1673- 5102.2014.01.012.
[22] CHEN X H, JIANG S, XU K, et al. Microsporogenesis and the forma- tion of male gametophytes in(Thunb.) Vahl. [J]. Acta Sci Nat Univ Nankai, 2007, 40(2): 79–82,104.陈旭辉, 江莎, 许珂, 等. 连翘的小孢子发生与雄配子体形成 [J]. 南开大学学报(自然科学版), 2007, 40(2): 79–82,104.
[23] KANG X Y. Study on mechanism of pollen abortion in Chinese white poplar (Carr.) [J]. Sci Silv Sin, 2001, 37(3): 35–39. doi: 10.11707/j.1001-7488.20010307.康向阳. 毛白杨花粉败育机制的研究 [J]. 林业科学, 2001, 37(3): 35–39. doi: 10.11707/j.1001-7488.20010307.
[24] ZHANG C H, TANG G Y, SUN Y Y, et al. Physiological characteristics of(L.) Jacq. seeds stress-resistance and influences on nature regeneration [J]. SW China J Agric Sci, 2010, 23(5): 1471– 1476. doi: 10.3969/j.issn.1001-4829.2010.05.019.张春华, 唐国勇, 孙永玉, 等. 车桑子种子抗逆生理学特性及其对天然更新的影响 [J]. 西南农业学报, 2010, 23(5): 1471–1476. doi: 10.3969/j.issn.1001-4829.2010.05.019.
[25] LIANG C L, LIU M J, ZHAO J. Research progress on plant seeds abortion [J]. Mol Plant Breed, 2005, 3(1): 117–122. doi: 10.3969/j.issn. 1672-416X.2005.01.020.梁春莉, 刘孟军, 赵锦. 植物种子败育研究进展 [J]. 分子植物育种, 2005, 3(1): 117–122. doi: 10.3969/j.issn.1672-416X.2005.01.020.
Microsporogenesis and Male Gametophyte Development of
LU Xue-jia, LIU Fang-yan, GAO Cheng-jie, LI Kun*
(Research Institute of Resource Insects, Chinese Academy of Forestry,Kunming 650224, China)
To understand the embryological characteristics ofin the dry and hot valley and the reasons for its low seed setting rate, microsporogenesis, male gametophyte development and pollen morphological characteristics ofwere studied with traditional paraffin sections and scanning electron microscopy. The results showed that the anther ofhad four pollen sac. The anther wall was composed of the epidermis, the endothecium, 2-3 middle layers, and tapetum (glandular tapetum) from the outside to the inside; at anther mature stage, the middle layer and tapetum denegerated and disappeared. The cytokinesis occurred simultaneously in a microspore mother cell; the tetrad was tetrahedral. The mature pollen was two-celled type. The mature pollen grain was nearly spherical with granular ornamentation on exine and 3 germ furrows which did not form syncolpate. Our researches showed that some abnormal phenomenon in male reproductive development might be one of the reasons of low seed setting rate.
; Microsporogenesis; Male gametophyte; Pollen; Morphological characteristics
10.11926/jtsb.3939
2018-05-04
2018-07-15
“十二五”国家科技计划课题(2015BAD07B0404);国家重点研发计划项目(2017YFC0505101)资助
This work was supported by the National Science and Technology Plan in the Twelfth Five-Year Period (Grant No. 2015BAD07B0404), and the National Key Research and Development Planning Program of China (Grant No. 2017YFC0505101).
卢雪佳(1993~ ),女,硕士研究生。研究方向为防护林。E-mail: 1771836520@qq.com
E-mail: caflikun@163.com
