井冈山鹿角杜鹃群落灌木层植物叶功能性状对海拔梯度的响应
2019-04-04向琳陈芳清耿梦娅王玉兵吕坤杨世林
向琳, 陈芳清*, 耿梦娅, 王玉兵, 吕坤, 杨世林
井冈山鹿角杜鹃群落灌木层植物叶功能性状对海拔梯度的响应
向琳1, 陈芳清1*, 耿梦娅1, 王玉兵1, 吕坤1, 杨世林2
(1. 湖北省三峡地区生态保护与治理国际联合研究中心,湖北 宜昌 443002;2. 湖北正江环保科技有限公司,湖北 宜昌 443002)
为揭示鹿角杜鹃()群落灌木层植物叶功能性状及其对环境变化的响应趋势,对分布于井冈山不同海拔梯度鹿角杜鹃群落灌木层植物的叶功能性状进行了研究。结果表明,海拔梯度对灌木植物的叶功能性状有显著影响。随海拔的升高,叶片的干物质含量(LDMC)、厚度(LT)、氮含量(LNC)、磷含量(LPC)呈显著上升趋势,比叶面积(SLA)和N/P呈显著下降趋势,而叶大小(LS)呈先上升后下降的变化趋势;灌木植物叶片的LDMC与SLA、LS呈负相关,与LT、LNC、LPC呈正相关;SLA与LT、LNC呈负相关;LS与LT呈负相关;LNC与LPC呈正相关;N/P与LPC呈负相关;环境因子对灌木植物叶功能性状有重要影响,除受海拔的影响外,LPC、N/P还受坡位的影响,LS、LNC则分别还受到坡向和坡度的影响。因此,井冈山地区鹿角杜鹃群落灌木层植物通过改变叶功能性状来适应海拔和其它环境因子的变化。
叶片;功能性状;海拔;干物质;鹿角杜鹃;井冈山
叶功能性状包括结构型性状和功能型性状,结构型性状有大小(leaf size, LS)、比叶面积(specific leaf area, SLA)、厚度(leaf thickness, LT)、干物质含量(leaf dry matter content, LDMC)等反映生物化学结构特征。功能型性状主要包括净光合速率、呼吸速率和气孔导度等与植物生理功能密切相关的生理生化参数。结构型性状稳定性相对较高,可以较好地反映植物为获得最大碳收获而采取的生存适应策略。不同植物种类的叶片性状对自然环境因子作用的响应常常存在着很大差异[9],同一地点内的物种共存其实是多物种生活策略的稳定组合[10]。开展同一群落类型各植物种类叶片功能性状随环境梯度变化的研究,对于了解和预测整个群落的资源利用及群落结构的动态具有重要的意义[6]。
井冈山位于南岭山地罗霄山脉中段,是江西境内分布的典型二元山体结构,地形多样,具有特殊的盆岭相间格架。复杂的地形地貌孕育了丰富的物种多样性,该地区是我国生物多样性分布的关键区域之一[11]。杜鹃属()植物是江西省分布最广、类型最多、面积最大的一类灌丛植物。鹿角杜鹃()作为杜鹃花属中垂直分布最宽的种类之一,对环境的变化具有较强的适应性。我们对分布于井冈山地区不同海拔的鹿角杜鹃群落进行了实地调查研究,测定了群落灌木优势种叶片的厚度、大小、干物质含量、比叶面积、单位质量的含氮量、含磷量和氮磷比等功能性状,对井冈山地区鹿角杜鹃群落灌木层优势种的叶功能性状特征、相互关系,叶功能性状随海拔梯度和环境因子的变化进行了分析与探讨,以揭示该群落类型对环境变化的响应趋势,为科学保护和利用鹿角杜鹃种质资源和生物多样性提供科学依据。
1 材料和方法
1.1 研究地概况
井冈山地处26°27′~26°40′ N,113°39′~114°23′ E,整个山体呈东北-西南走向,海拔为202~1 841 m,相对高差较大。土壤以山地黄壤为主,其他类型土壤随海拔增加呈现规律的垂直带谱分布。该区位于南亚热带与中亚热带的过渡地带,为亚热带湿润季风气候。年平均气温为14℃~17℃,年降雨量1 856.2 mm。典型植被为常绿阔叶林,拥有灌木林、山顶矮林、针叶林、针阔叶混交林、落叶阔叶林等多种森林群落类型,森林覆盖率高达81.2%[11–12]。以杜鹃属植物为建群种的群落主要分布在较高海拔的山地,其中鹿角杜鹃广泛生长在井冈山全山300~1 200 m处,群落环境具有湿度高、温度低、营养资源丰富的特点。本研究所样地的土壤类型为山地黄壤或黄棕壤,pH 4.9~5.2,土壤含水量及N、P含量在样地间有较大差异(表1)。
1.2 野外调查和样品采集
以井冈山地区海拔600~1500 m为研究区域,在低海拔、中海拔和高海拔处挑选具有代表性的鹿角杜鹃群落,各设置3个20 m×20 m的样地,每个样地随机设置3个5 m×5 m的样方,测量统计样方内灌木层各物种的株数、高度、盖度等,并且测量样地的经纬度、含水量、海拔等环境因子(表1)。低海拔3个样地的海拔分别为647、662和688 m,灌木以鹿角杜鹃为主,伴生马银花()、大叶早樱()、毛肉黄楠()和枳椇(),草本层不发达,腐殖质丰富;中海拔3个样地的海拔分别为820、845和860 m,鹿角杜鹃为优势灌丛,伴生长蕊杜鹃()、猴头杜鹃(),生长稀疏,草本极稀,苔藓层覆盖度高;高海拔3个样地的海拔分别为1 340、1 370和1 394 m,鹿角杜鹃散生分布,常见长蕊杜鹃、满山红()、交让木(),地面裸露黄棕土并覆盖大量枯枝落叶。3个样地均无高大乔木,这里将高度不足5 m的乔木树种均视为灌木[13]。
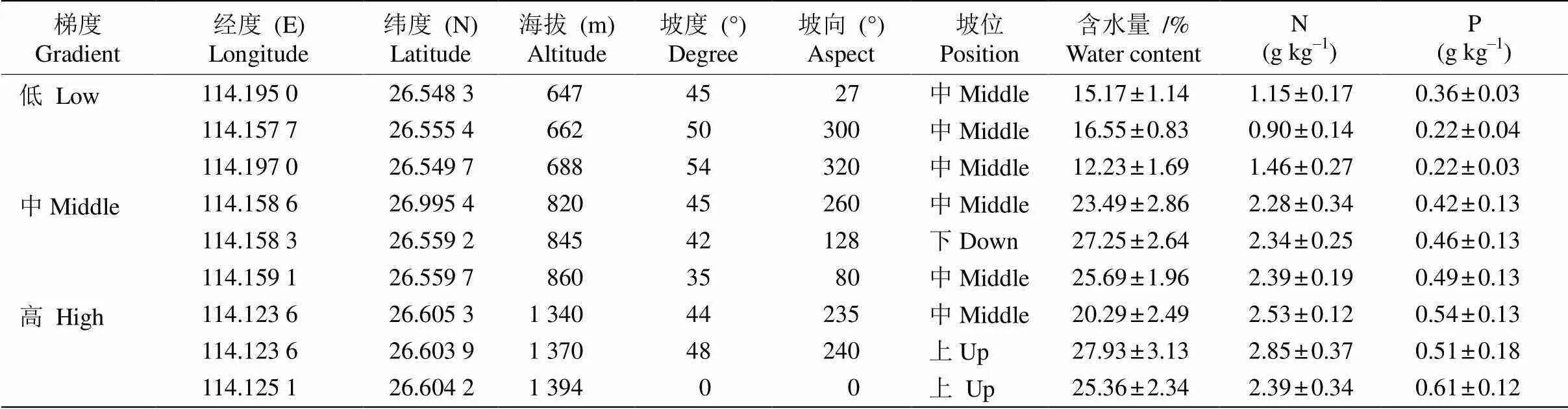
表1 不同海拔群落的基本特征
在每个样方内选取重要值大于0.1的优势物种[14],每种植物随机选取5株生长成熟、长势良好的植株,每株植物随机挑选分布于5个方向、完全展开的10片叶片,共50片叶片。将所有的叶片放置于两片湿润滤纸之间,封入自封袋,带回室内测量各性状指标[14]。各群落中的草本层均较为稀疏,故没有进行草本层植物叶功能性状的调查。
1.3 叶功能性状的测定
从采集的50片叶中选取20片在5℃避光下放置12 h后擦干叶表面水分,用精度为0.001的电子天平称量叶片的饱和鲜重。然后用WinRHIZO根系分析系统扫描叶片测定大小(LS),同时在叶片主脉上均匀选择3个点,使用游标卡尺(精度为0.02 mm)测量每个点旁约0.25 cm处的厚度,取平均值作为叶片的厚度(LT)[15]。最后将叶片放进80℃烘箱内烘24 h,取出称量叶片干重。计算比叶面积(SLA, m2g–1)=叶片面积(m2)/叶片干重(kg)。叶干物质含量(LDMC, g kg–1)=叶片干重(g)/叶片饱和鲜重(kg)[1]。剩余的30片在65℃下烘干至恒重,用植物粉碎机粉碎过筛,用San++连续流动分析仪测定全氮(leaf nitrogen content, LNC)、全磷(leaf phosphorus content,LPC)含量,LNC (g kg–1)=叶片全氮(g)/叶片干重(kg); LPC (g kg–1)=叶片全磷(g)/叶片干重(kg); N/P= LNC/LPC。重复3次,取平均值。
1.4 数据分析和处理
灌木层各物种重要值=(相对密度+相对盖度+相对频度)/3。9个样地的优势种共有26种。
将同一海拔所有优势种的叶性状值按其重要值进行加权得到各海拔的叶功能性状值,用Excel 2012完成计算。
2.消瘦 随着呕吐的加剧,由于奶水摄入不足,体重起初不增,之后迅速下降,尿量明显减少,数日排便1次,量少且质硬,偶有排出棕绿色便,被称为饥饿性粪便。由于营养不良、脱水,婴儿明显消瘦,皮肤松弛有皱纹,皮下脂肪减少,精神抑郁呈苦恼面容。发病初期呕吐丧失大量胃酸,可引起碱中毒,呼吸变浅而慢,并可有喉痉挛及手足搐搦等症状;后期脱水严重,肾功能低下,酸性代谢产物潴留体内,部分碱性物质被中和,故很少有明显碱中毒者。严重营养不良的晚期病例已难以见到。
利用单因素方差分析法(One-Way ANOVA)分析不同海拔梯度间群落各叶性状的差异显著性, 并进行叶功能性状间及性状与地形因子和土壤理化性质间的Pearson相关性分析。所有性状和海拔、坡度、土壤性质数据均以现场测量值表示,且具有正态分布特征。坡位分别以1、2、3代表上坡位、中坡位和下坡位;坡向数据以朝东为起点(即为0°)顺时针旋转,每45°为1个区间:1表示北坡(247.5°~ 292.5°),2表示东北坡(292.5°~337.5°),3表示西北坡(202.5°~247.5°),4表示东坡(337.5°~22.5°),5表示西坡(167.5°~202.5°),6表示东南坡(22.5°~ 67.5°),7表示西南坡(112.5°~167.5°),8表示南坡(67.5°~112.5°),数字越大,代表越向阳越干热[18]。以上分析均利用SPSS 19.0进行。
选取LDMC、SLA、LT、LS、LNC、LPC和N/P等7个叶功能性状为因变量,以海拔、坡度、坡位、坡向4个地形因子和土壤N、P、含水量共7个环境因子为自变量,在SPSS 19.0软件中利用逐步回归建立线性回归模型。标准偏回归系数越大的环境因子,表示对叶功能性状的影响越大。
2 结果和分析
2.1 不同海拔的叶片功能性状比较
海拔梯度对鹿角杜鹃群落植物叶片的LDMC、SLA、LT、LS、LNC、LPC和N/P等功能性状均有显著影响(<0.05)(图1)。LDMC、LT、LNC和LPC随海拔上升而增加,SLA、N/P随海拔增加而减少,LS呈先增加后减小的趋势。从低海拔到高海拔群落的LDMC均值分别为428.34、447.48、468.88 g kg–1,LT分别为0.38、0.41和0.46 mm, 高海拔群落的LDMC和LT分别比低海拔的增加9.46%和21.05%;从低海拔到高海拔的SLA分别为43.03、28.97和23.86 m2kg–1,海拔梯度间均有显著差异; N/P从低海拔到高海拔分别为2.92、1.58和1.54, 低海拔与中海拔和高海拔间有显著差异,中海拔与高海拔间差异不明显;LNC从低海拔到高海拔分别为5.01、5.15和5.48 g kg–1,低海拔和中海拔间不存在显著差异,但两者与高海拔间有显著差异;LPC和LS从低海拔到高海拔分别为1.73、3.29、3.56 g kg–1和63.35、64.74、42.13 cm2,且低海拔与中海拔和高海拔间有显著差异,但中海拔与高海拔间差异不显著。
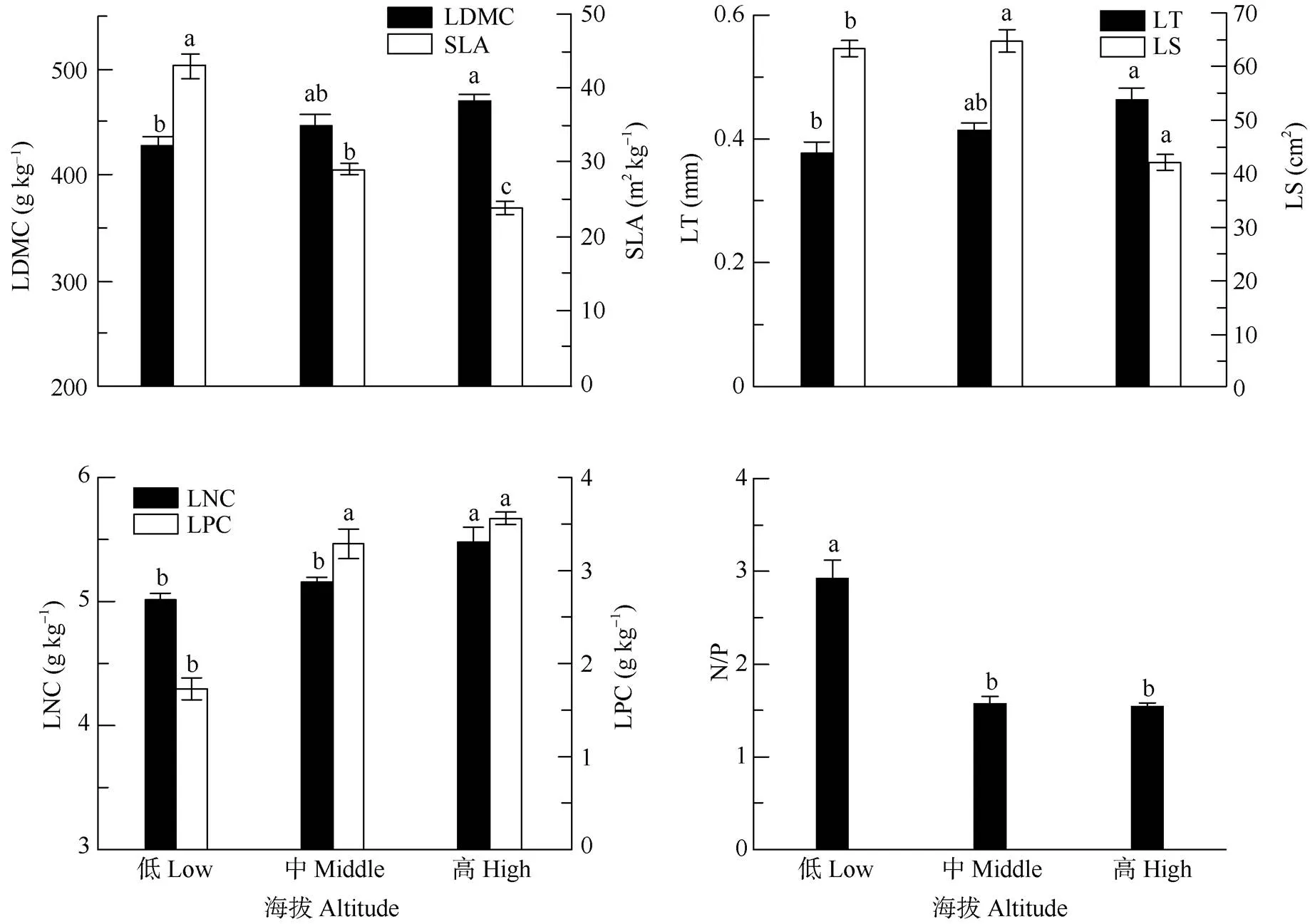
图1 海拔梯度对鹿角杜鹃群落植物叶功能性状的影响。柱上不同字母表示差异显著(P<0.05); LDMC: 干物质含量; SLA: 比叶面积; LT: 厚度; LS: 大小; LNC: 氮含量; LPC: 磷含量。下表同。
2.2 相关性分析
Pearson相关检验表明(表2),LDMC与LT呈极显著正相关,与LNC、LPC呈显著正相关,与SLA、LS呈显著负相关;SLA与LS呈极显著正相关,与LNC、LT呈显著负相关;LT与LNC、LPC呈显著正相关,与LS、N/P呈显著负相关;LS与LNC呈显著负相关;LNC与LPC呈显著正相关; LPC与N/P呈极显著负相关。
2.3 叶功能性状与环境因子间的相关性
从表3可见,LDMC与土壤P含量的相关系数最大,呈极显著正相关,与海拔和土壤N含量呈显著正相关,与其他环境因子则无显著相关性;SLA与海拔呈极显著负相关,与其他环境因子无显著相关性;LT与海拔和土壤N含量呈极显著正相关,与土壤N含量的相关系数最大,与土壤P含量呈显著正相关,与其他环境因子无显著相关性;LS与海拔呈极显著负相关,与其他环境因子均无显著相关性;N/P与海拔、土壤N含量、含水量均呈极显著负相关,与土壤N含量的相关系数最大,与土壤P含量呈显著负相关,与坡度、坡位、坡向无显著相关性;LNC与海拔和土壤P含量呈极显著正相关,与海拔的相关系数最大,与坡度、坡位和土壤N含量呈显著负相关,与坡向和土壤含水量无显著相关性;LPC与土壤N的相关系数最大,与土壤N、P含量、含水量均呈极显著正相关,与海拔呈显著正相关,与坡度、坡位、坡向无显著相关性;N/P与海拔、土壤N、P含量和含水量呈极显著负相关,与土壤N含量的相关系数最大,与土壤P含量呈显著负相关,与其他环境因子无显著相关性。

表2 叶功能性状的Pearson相关性分析
*:<0.05;**:<0.01
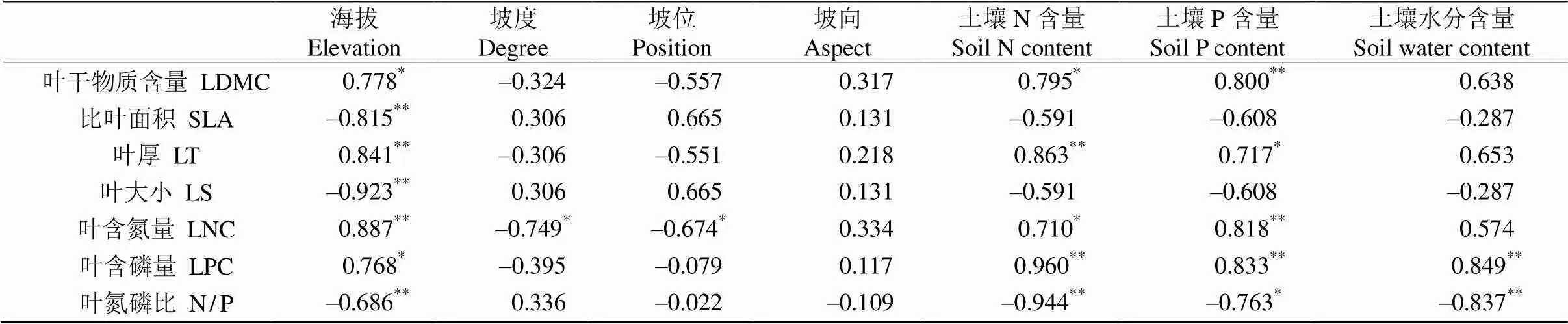
表3 叶功能性状与环境因子间的Pearson相关性
*:<0.05;**:<0.01
2.4 叶功能性状与环境因子的逐步回归分析
叶功能性状与环境因子的逐步回归分析表明, 不同环境因子对叶功能性状的影响力有差异(表4)。所有的叶功能性状均受到海拔的影响最大,海拔对LDMC、SLA、LT、LS、LNC、LPC、N/P的标准偏回归系数分别为0.778、–0.815、0.841、–0.969、0.681、1.153和–1.121。除海拔外,LPC、N/P主要受到坡位的影响,LS主要受到坡向的影响,LNC主要受到坡度的影响。LDMC、LS和LNC主要受到土壤P含量的影响,标准偏回归系数分别为0.800、–0.608和0.818; LT、LPC、N/P则主要受到土壤N含量的影响,标准偏回归系数分别为0.795、0.960和–0.944;LS受土壤因子的影响不显著;SLA未明显受土壤因子的影响。
3 讨论
3.1 群落植物叶功能性状对海拔梯度的响应特征
海拔是重要的地形因子,太阳辐射强度、土壤养分、降水量等水热环境因子均随着海拔的变化而发生变化。张继平等[12]对井冈山自然保护区的土壤进行研究, 表明土壤全氮全磷含量均随着海拔升高呈增加趋势。而海拔梯度变化引起水热的重新分配以及土壤环境因子的变化会引起植物叶片性状发生复杂的适应性变化,并使植物形成不同功能性状组合的生活策略[16]。
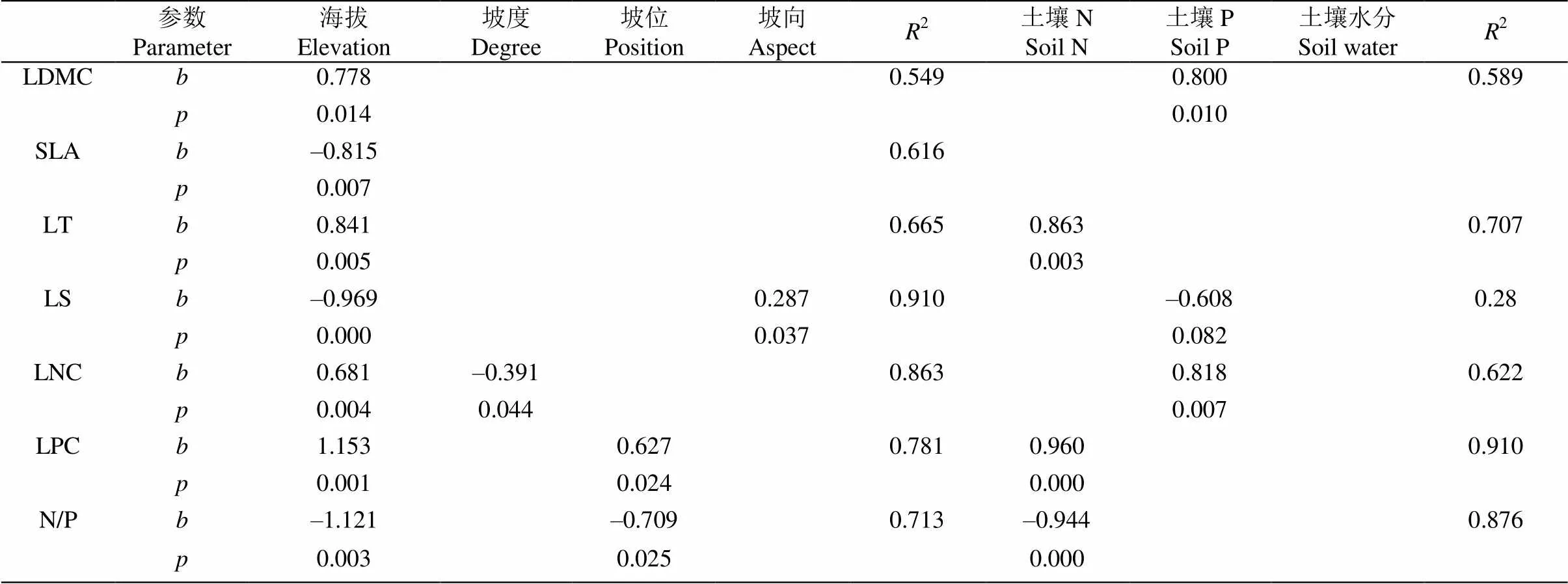
表4 叶功能性状与环境因子间的逐步回归
: 标准偏回归系数
: Standard patrial regression coefficient
SLA和LDMC主要反映了植物的资源利用效率以及对养分的保存能力,与环境的变化联系密切,是研究群落间差异的首要性状[14]。井冈山地区鹿角杜鹃群落的LDMC随海拔升高而增加,SLA则减小。杨锐等报道云南元谋干热河谷植物SLA随海拔升高而减小[17]。李宗杰等[18]调查了摩天岭北坡高、低海拔区的森林群落,均表现出SLA随海拔升高而降低的趋势。但罗璐[6]等报道神农架4种典型森林的SLA随海拔升高而增加,这是因为高海拔森林群落中落叶树种增多,落叶树种的SLA往往大于常绿树种。Zhang等[19]也认为SLA随海拔升高而增加。宝乐等[14]对东灵山地区不同森林群落和栾志慧等[20]对长白山牛皮杜鹃()的LDMC与海拔梯度关系的研究结果与本文一致。SLA随海拔升高呈现下降趋势,植物叶片往往会通过细胞大小和细胞间隙的减少来提高其御寒能力因而导致SLA的减小[21],这可能是井冈山鹿角杜鹃群落灌木层植物对高海拔低温的适应性对策。
LS往往与植物的光截取和碳获取能力有关,因此在不同生境间有很大的变化[8]。很多研究表明, LS随海拔升高而下降[9,16],本研究表明,LS随海拔升高呈先增加后下降的趋势,总体呈现下降趋势。这可能是因为海拔升高气温降低,在低温环境下叶片接受光能获取能量的效率降低,叶片需要通过减小叶面积来减少呼吸和蒸腾消耗[8]。在低海拔地区由于人类干扰程度较大,LS略小于干扰力度相对较小的中海拔地区。LT通常与养分的利用有关[15]。有研究表明,LT随海拔升高而下降[6]。但本研究中LT随海拔升高而增加,与Li等[22]的研究结果相同。这是由于本研究区域海拔高差相对较小,降水量变化不明显,环境水分的影响相对较小,而海拔的升高引起温度的降低却比较明显,且降温能增加叶片的密度和厚度[23]。
氮对植物的生长发育和生产力具有重要的影响,同时也影响植物对干旱的适应[8]。单位叶面积的氮含量普遍随海拔升高而增加[6,24],而单位叶质量的LNC对海拔的响应趋势不尽相同。Craine等[9]报道随海拔升高,叶和根的氮含量降低。王晶媛等[25]报道随海拔的增加,LNC和叶宽均呈先下降后上升的二次曲线变化趋势。但祁建等[26]和罗璐等[6]的研究表明LNC、LPC随海拔的升高呈增加趋势。Xu等[27]认为海拔与LNC呈线性显著正相关, LPC与LNC存在着动态平衡,具有协同作用,变化往往具有一致性[18]。本研究中,LNC和LPC均随海拔升高而增加,存在显著正相关,与土壤氮、磷含量变化趋势一致,这与顾大形等[28]关于LNC、LPC与土壤氮、磷含量呈正相关的结论一致。植物叶片的N/P可以作为判断环境对植物生长的养分供应状况的指标[29],本研究中叶片N/P沿海拔梯度的变化趋势与LNC、LPC的变化趋势完全相反,随海拔升高呈下降趋势,这可能与温度的降低有关, 温度的升高往往会导致N/P的升高[30],这与杨阔等[31]的研究结论一致。在井冈山3个海拔梯度中鹿角杜鹃群落灌木层植物叶片N/P含量偏低,土壤N或P含量的增加预计不可能提高整个群落的物种丰富度[32]。而低N/P的群落往往采取快速获取外部资源的生存策略[33],容易受到外界干扰的影响。随着人为干扰的日益增加,群落一旦破坏,往往难以恢复,在高海拔地区应该加以保护,以经营风景林为主, 而在低海拔发展人工林的同时,保留这些具有药用价值的灌木,在经济发展的同时注重生物多样性的保护。
3.2 叶功能性状间的相关关系
植物功能性状间紧密的相关关系是普遍存在的,这是由于物种间在生长发育及形成适应性机制的过程中相互作用,表现出相对应的生态策略,使群落内各物种都能够具有相应的生态位[20]。很多研究表明,植物的SLA与LDMC呈现显著负相关关系[28,30,34],SLA与LT呈显著的负相关关系[18], LDMC与LS呈负相关关系,而SLA与LS呈正相关关系[35],上述研究结论均与本研究结果一致。
LDMC与LT的关系较为复杂。本研究中, LDMC与LT呈极显著正相关关系,与罗璐等[6]、李宗杰等[18]、Ackerly等[36]的研究结论一致。但胡耀升等[35]和杨锐等[17]则报道LDMC与LT呈负相关。已有大量研究阐明,LDMC越大表明叶组织密度也越高,叶则越坚韧,其厚度越大,越能抵抗威胁生长的物理因子[37]。曾小平等[38]和刘贵峰等[35]的研究表明LS与LT呈极显著正相关关系,而本研究表明LS与LT呈负相关关系。LS和LT受环境因素影响较大[39],在诸多环境因子影响下,群落中植物叶片的大小和厚度形成最适宜生长的生存策略,LS和LT负相关并不表示该群落的LS和LT在植物吸收或储存过程中存在拮抗作用,而是沿环境梯度形成对策的变化趋势刚好不同。也有研究表明叶片密度和LT通常是独立的,与叶片的其他功能性状的相关性并不明显[40]。
本研究结果还表明,井冈山地区鹿角杜鹃群落叶功能性状中SLA与LNC呈负相关关系,LDMC与LNC、LPC呈正相关,这与罗璐等[6]、李宏伟等[41]、宝乐等[14]的研究结果不一致。这可能是由于环境梯度的影响及土壤氮、磷含量引发植物实现自我调节来适应环境的变化,与一般的分配策略不同,形成了与环境相对应的生存策略,使得LNC与SLA呈现负相关关系。N/P与LPC呈极显著负相关,这与Chen等[30]、Güsewell等[40]和宋彦涛等[42]的研究结论一致。
3.3 叶功能性状与环境因子的关系
海拔、坡位、坡度、坡向等地形因子主要是通过改变太阳辐射强度、温度、降水量和土壤条件间接影响植物的生长和分布。在山地生境中,除了海拔等因子会影响植物的性状,坡位、坡向、坡度等也是重要的影响因素[26]。海拔的变化往往会引起温度和水分条件的差异,随海拔的升高,植物叶片形态的可塑性往往也会发生变化[23],通过对井冈山地区鹿角杜鹃群落优势种植物叶功能性状与地形因子的逐步回归分析,表明各功能性状均受到海拔的影响最大,这可能是因为井冈山地区气候温暖,生长环境优越,最主要的外界干扰就是引起温度和水分变化最大的海拔。除海拔因子外,LPC、N/P主要还受到坡位的影响,坡向和坡度也分别对LS和LNC有很大的影响。在山地环境中,土壤深度、质地和矿物质含量都会受到坡位和坡度的影响,上坡位往往光照较强,温度较高,土壤营养元素缺失[26]。宝乐等[14]对东灵山主要森林群落优势种叶功能性状的研究表明坡位是LPC的首要影响因子。LS的形成机制与适应性策略往往与叶能量的平衡变化有关[43]。Ackerly等[35]对美国加利福尼亚海岸群落灌木树种的研究表明,在北坡面没有小叶树种,群落水平上的LS变化与日照强度有关。
植物的生存往往也受到土壤因子的影响,土壤条件的改变影响了植物对营养元素的获取策略,引起植物性状的变异,形成相适应的生存方式[33]。大多数叶性状均与土壤氮、磷和含水量具有显著相关性。胡耀升等[44]报道土壤氮含量是SLA的显著影响因子;Poorter等[45]的研究表明,植物LMA与温度、土壤磷含量关系密切。刘旻霞等[33]和丁佳等[46]认为土壤pH是影响植物性状变异的因子之一,能够通过影响植物对土壤磷的利用效率来影响植物的光合效率,进而影响到株高以及叶大小。本研究与以上研究结果不尽相同,井冈山鹿角杜鹃群落的LT、LPC、N/P主要受到土壤氮含量的影响,土壤磷含量是LDMC、LNC的首要影响因子,SLA并没有受到明显的土壤因子的影响。在井冈山地区土壤磷含量相对比较丰富,不是制约植物生长的首要限制性因子。LNC与土壤氮、磷含量具有显著相关性,LPC与N/P与土壤氮、磷和含水量均具有显著相关性,土壤养分的全氮、全磷和含水量在一定程度上影响了LPC、LNC。逐步回归分析结果表明, N/P主要受到土壤氮的影响。与顾大形等[28]对四季竹()叶片的研究结论一致, 这可能是由于土壤较低的氮含量限制了叶片对P的吸收,LPC与土壤N的极显著相关性和LPC与N/P的极显著相关性验证了这一观点。
致谢 本研究中官守鹏、谭向前和张行参加了野外调查,在此表示感谢!
[1] Cornelissen J H C, Lavorel S, Garnier E, et al. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide [J]. Aust J Bot, 2003, 51(4): 335–380. doi: 10.1071/ BT02124.
[2] Bernard-Verdier M, Navas M L, Vellend M, et al. Com- munity assembly along a soil depth gradient: Contrasting patterns of plant trait convergence and divergence in a Mediterranean rangeland [J]. J Ecol, 2012, 100(6): 1422–1433. doi: 10.1111/1365-2745.12003.
[3] KIMBALL S, FUNK J L, SPASOJEVIC M J, et al. Can functional traits predict plant community response to global change? [J]. Eco- sphere, 2016, 7(12): e01602. doi: 10.1002/ecs2.1602.
[4] LIU X J, MA K P. Plant functional traits-concepts, applications and future directions [J]. Sci Sin Vit, 2015, 45(4): 325–339. doi: 10.1360/ N052014-00244.刘晓娟, 马克平. 植物功能性状研究进展[J]. 中国科学: 生命科学, 2015, 45(4): 325–339. doi: 10.1360/N052014-00244.
[5] LIU M X. Studies on physiological and leaf morphological traits for photosynthesis on different slopes in a subalpine meadow [J]. Acta Ecol Sin, 2017, 37(24): 8526–8536. doi: 10.5846/stxb201610132072.刘旻霞. 亚高寒草甸不同坡向植物光合生理和叶片形态差异[J]. 生态学报, 2017, 37(24): 8526–8536. doi: 10.5846/stxb201610132072.
[6] LUO L, SHEN G Z, XIE Z Q, et al. Leaf functional traits of four typical forests along the altitudinal gradients in Mt. Shennongjia [J]. Acta Ecol Sin, 2011, 31(21): 6420–6428. 罗璐, 申国珍, 谢宗强, 等. 神农架海拔梯度上4种典型森林的乔木叶片功能性状特征[J]. 生态学报, 2011, 31(21): 6420–6428.
[7] ZHAO L C, ZHAO C Z, CHEN J, et al. Photosynthetic characteristics and twig-leaf traits of different densities ofin Qinwangchuan wetland [J]. Acta Ecol Sin, 2018, 38(5): 1722–1730. doi: 10.5846/stxb201702230296.赵连春, 赵成章, 陈静, 等. 秦王川湿地不同密度柽柳枝-叶性状及其光合特性[J]. 生态学报, 2018, 38(5): 1722–1730. doi: 10.5846/ stxb201702230296.
[8] WANG C S, WANG S P. A review of research on responses of leaf traits to climate change [J]. Chin J Plant Ecol, 2015, 39(2): 206–216. doi: 10.17521/cjpe.2015.0020.王常顺, 汪诗平. 植物叶片性状对气候变化的响应研究进展[J]. 植物生态学报, 2015, 39(2): 206–216. doi: 10.17521/cjpe.2015.0020.
[9] CRAINE J M, LEE W G. Covariation in leaf and root traits for native and non-native grasses along an altitudinal gradient in New Zealand [J]. Oecologia, 2003, 134(4): 471–478. doi: 10.1007/s00442-002-1155-6.
[10] WESTOBY M, FALSTER D S, MOLES A T, et al. Plant ecological strategies: Some leading dimensions of variation between species [J]. Annu Rev Ecol Syst, 2002, 33(1): 125–159. doi: 10.1146/annurev.ecol sys.33.010802.150452.
[11] CHEN B M, LIN Z G, LI Z, et al. Ecosystem diversity in Jinggangshan area, China [J]. Acta Ecol Sin, 2012, 32(20): 6326–6333. doi: 10.5846/ stxb201206120848.陈宝明, 林真光, 李贞, 等. 中国井冈山生态系统多样性[J]. 生态学报, 2012, 32(20): 6326–6333. doi: 10.5846/stxb201206120848.
[12] ZHANG J P, ZHANG L B, WANG F Y, et al. Spatial variation of soil nutrient contents in the Jinggangshan National Nature Reserve [J]. Soils, 2014, 46(2): 262–268. doi: 10.13758/j.cnki.tr.2014.02.011.张继平, 张林波, 王风玉, 等. 井冈山国家级自然保护区森林土壤养分含量的空间变化[J]. 土壤, 2014, 46(2): 262–268. doi: 10. 13758/j.cnki.tr.2014.02.011.
[13] CUI D F. Plant Taxonomy [M]. Beijing: China Agriculture Press, 1994: 34. 崔大方. 植物分类学[M]. 北京: 中国农业出版社, 1994: 34.
[14] BAO L, LIU Y H. Comparison of leaf functional traits in different forest communities in Mt. Dongling of Beijing [J]. Acta Ecol Sin, 2009, 29(7): 3692–3703. doi: 10.3321/j.issn:1000-0933.2009.07.030.宝乐, 刘艳红. 东灵山地区不同森林群落叶功能性状比较[J]. 生态学报, 2009, 29(7): 3692–3703. doi: 10.3321/j.issn:1000-0933.2009. 07.030.
[15] LIU J H, ZENG D H, LEE D K. Leaf traits and their interrelationships of main plant species in southeast Horqin sandy land [J]. Chin J Ecol, 2006, 25(8): 921–925. doi: 10.13292/j.1000-4890.2006.0175.刘金环, 曾德慧, LEE D K. 科尔沁沙地东南部地区主要植物叶片性状及其相互关系[J]. 生态学杂志, 2006, 25(8): 921–925. doi: 10. 13292/j.1000-4890.2006.0175.
[16] JIANG A P, JIANG J M, LIU J. Responses of leaf traits of(Hemsl.) Hemsl. along an altitudinal gradient [J]. Chin J Ecol, 2016, 35(6): 1467–1474. doi: 10.13292/j.1000-4890.201606.017.蒋艾平, 姜景民, 刘军. 檫木叶片性状沿海拔梯度的响应特征[J]. 生态学杂志, 2016, 35(6): 1467–1474. doi: 10.13292/j.1000-4890. 201606.017.
[17] YANG R, ZHANG B R, WANG L L, et al. The response of plant functional traits’ group to gradients of altitude in Dry-hot Valley of Yuanmou [J]. Ecol Environ Sci, 2015, 24(1): 49–56. doi: 10.16258/j. cnki.1674-5906.2015.01.008.杨锐, 张博睿, 王玲玲, 等. 元谋干热河谷植物功能性状组合的海拔梯度响应[J]. 生态环境学报, 2015, 24(1): 49–56. doi: 10.16258/j. cnki.1674-5906.2015.01.008.
[18] LI Z J, TIAN Q, SONG L L. Variation and correlation of leaf traits in woody plants in the north-facing slope of Motianling, Gansu, China [J]. J Desert Res, 2018, 38(1): 149–156. 李宗杰, 田青, 宋玲玲. 甘肃省摩天岭北坡木本植物叶性状变异及关联[J]. 中国沙漠, 2018, 38(1): 149–156.
[19] ZHANG L, LUO T X, LIU X S, et al. Altitudinal variation in leaf construction cost and energy content of[J]. Acta Oecol, 2012, 43: 72–79. doi: 10.1016/j.actao.2012.05.011.
[20] LUAN Z H, SHAO D K, YANG L J, et al. Comparative analysis on adaptation characteristic ofPall. leaves in different altitudes of Changbai Mountain [J]. N Hort, 2013(19): 80–83. 栾志慧, 邵殿坤, 杨丽娟, 等. 不同海拔长白山牛皮杜鹃叶片适应性结构的对比分析[J]. 北方园艺, 2013(19): 80–83.
[21] CORDELL S, GOLDSTEIN G, MUELLER-DOMBOIS D, et al. Physiological and morphological variation in, a dominant hawaiian tree species, along an altitudinal gradient: The role of phenotypic plasticity [J]. Oecologia, 1998, 113(2): 188–196. doi: 10.1007/s004420050367.
[22] LI F L, BAO W K. Elevational trends in leaf size ofin the arid Minjiang River valley, SW China [J]. J Arid Environ, 2014, 108: 1–9. doi: 10.1016/j.jaridenv.2014.04.011.
[23] YANG D M, ZHANG J J, ZHOU D, et al. Leaf and twig functional traits of woody plants and their relationships with environmental change: A review [J]. Chin J Ecol, 2012, 31(3): 702–713. doi: 10.13292/ j.1000-4890.2012.0152.杨冬梅, 章佳佳, 周丹, 等. 木本植物茎叶功能性状及其关系随环境变化的研究进展[J]. 生态学杂志, 2012, 31(3): 702–713. doi: 10. 13292/j.1000-4890.2012.0152.
[24] HIKOSAKA K, NAGAMATSU D, ISHII H S, et al. Photosynthesis- nitrogen relationships in species at different altitudes on Mount Kinabalu, Malaysia [J]. Ecol Res, 2002, 17(3): 305–313. doi: 10.1046/j. 1440-1703.2002.00490.x.
[25] WANG J Y, ZHANG H, YU M K, et al. Response of leaf traits ofenvironmental factors at regional scale [J]. Ecol Environ Sci, 2017, 26(5): 754–762. doi: 10.16258/j.cnki.1674-5906. 2017.03.005.王晶媛, 张慧, 虞木奎, 等. 区域尺度上麻栎叶片性状对环境因子的响应规律[J]. 生态环境学报, 2017, 26(5): 754–762. doi: 10.16258/ j.cnki.1674-5906.2017.03.005.
[26] QI J, MA K M, ZHANG Y X. The altitudinal variation of leaf traits ofand associated environmental explanations [J]. Acta Ecol Sin, 2007, 27(3): 930–937. doi: 10.3321/j.issn:1000-0933. 2007.03.013.祁建, 马克明, 张育新. 辽东栎()叶特性沿海拔梯度的变化及其环境解释[J]. 生态学报, 2007, 27(3): 930–937. doi: 10.3321/j.issn:1000-0933.2007.03.013.
[27] XU P B, ZHANG X W, ZHAO C M, et al. Foliar responses ofFranch. to altitude in the Taibai Mountain, China [J]. Pol J Ecol, 2014, 62(3): 479–490. doi: 10.3161/104.062.0309.
[28] GU D X, CHEN S L, HUANG Y Q. Effects of soil nitrogen and pho- sphonium on leaf nitrogen and phosphonium stoichiometric charac- teristics and chlorophyll content of[J]. Chin J Plant Ecol, 2011, 35(12): 1219–1225. doi: 10.3724/SP.J.1258.2011. 01219.顾大形, 陈双林, 黄玉清. 土壤氮磷对四季竹叶片氮磷化学计量特征和叶绿素含量的影响[J]. 植物生态学报, 2011, 35(12): 1219– 1225. doi: 10.3724/SP.J.1258.2011.01219.
[29] AERTS R, CHAPIN F S III. The mineral nutrition of wild plants revisited: A re-evaluation of processes and patterns [J]. Adv Ecol Res, 1999, 30: 1–67. doi: 10.1016/S0065-2504(08)60016-1.
[30] CHEN Y H, HAN W X, TANG L Y, et al. Leaf nitrogen and pho- sphorus concentrations of woody plants differ in responses to climate, soil and plant growth form [J]. Ecography, 2013, 36(2): 178–184. doi: 10.1111/j.1600-0587.2011.06833.x.
[31] YANG K, HUANG J H, DONG D, et al. Canopy leaf N and P stoichiometry in grassland communities of Qinghai-Tibetan Plateau, China [J]. Chin J Plant Ecol, 2010, 34(1): 17–22. doi: 10.3773/j.issn. 1005-264x.2010.01.004.杨阔, 黄建辉, 董丹, 等. 青藏高原草地植物群落冠层叶片氮磷化学计量学分析[J]. 植物生态学报, 2010, 34(1): 17–22. doi: 10.3773/j. issn.1005-264x.2010.01.004.
[32] GÜSEWELL S, BAILEY K M, ROEM W J, et al. Nutrient limitation and botanical diversity in wetlands: Can fertilisation raise species richness? [J]. Oikos, 2005, 109(1): 71–80. doi: 10.1111/j.0030-1299. 2005.13587.x.
[33] LIU M X, MA J Z. Responses of plant functional traits and soil factors to slope aspect in alpine meadow of South Gansu, Northwest China [J]. Chin J Appl Ecol, 2012, 23(12): 3295–3300. doi: 10.13287/j.1001- 9332.2012.0410.刘旻霞, 马建祖. 甘南高寒草甸植物功能性状和土壤因子对坡向的响应[J]. 应用生态学报, 2012, 23(12): 3295–3300. doi: 10.13287/j. 1001-9332.2012.0410.
[34] WRIGHT I J, REICH P B, WESTOBY M. Strategy shifts in leaf physiology, structure and nutrient content between species of high- and low-rainfall and high- and low-nutrient habitats [J]. Funct Ecol, 2001, 15(4): 423–434. doi: 10.1046/j.0269-8463.2001.00542.x.
[35] LIU G F, LIU Y P, BAIYILA D F, et al. Leaf traits of dominant plants of main forest communities in Daqinggou Nature Reserve [J]. Acta Ecol Sin, 2017, 37(14): 4646–4655. doi: 10.5846/stxb201603290575.刘贵峰, 刘玉平, 达福白乙拉, 等. 大青沟自然保护区主要森林群落优势种的叶性状[J]. 生态学报, 2017, 37(14): 4646–4655. doi: 10. 5846/stxb201603290575.
[36] ACKERLY D, KNIGHT C, WEISS S, et al. Leaf size, specific leaf area and microhabitat distribution of chaparral woody plants: Contrasting patterns in species level and community level analyses [J]. Oecologia, 2002, 130(3): 449–457. doi: 10.1007/s004420100805.
[37] XIE Y J. The characteristics of 20 dominant plant functional traits in evergreen broad-leaved forest in Daming Mountain Nature Reserve, Guangxi [D]. Nanning: Guangxi University, 2013: 46–47. 谢益君. 广西大明山常绿阔叶林20种优势植物的功能性状特征[D]. 南宁: 广西大学, 2013: 46–47.
[38] ZENG X P, ZHAO P, CAI X A, et al. Shade-tolerance of 25 low subtropical plants [J]. J Beijing For Univ, 2006, 28(4): 88–95. doi: 10. 13332/j.1000-1522.2006.04.017.曾小平, 赵平, 蔡锡安, 等. 25种南亚热带植物耐阴性的初步研究[J]. 北京林业大学学报, 2006, 28(4): 88–95. doi: 10.13332/j.1000- 1522.2006.04.017.
[39] HUANG M, JI J J. The spatial-temporal distribution of leaf area index in China: A comparison between ecosystem modeling and remote sensing reversion [J]. Acta Ecol Sin, 2010, 30(11): 3057–3064. 黄玫, 季劲钧. 中国区域植被叶面积指数时空分布——机理模型模拟与遥感反演比较[J]. 生态学报, 2010, 30(11): 3057–3064.
[40] GÜSEWELL S, KOERSELMAN W. Variation in nitrogen and pho- sphorus concentrations of wetland plants [J]. Perspect Plant Ecol, 2002, 5(1): 37–61. doi: 10.1078/1433-8319-0000022.
[41] LI H W, WANG X A, GUO H, et al. Leaf functional traits of different forest communities in Ziwuling Mountains of Loess Plateau [J]. Chin J Ecol, 2012, 31(3): 544–550. doi: 10.13292/j.1000-4890.2012.0146.李宏伟, 王孝安, 郭华, 等. 黄土高原子午岭不同森林群落叶功能性状[J]. 生态学杂志, 2012, 31(3): 544–550. doi: 10.13292/j.1000- 4890.2012.0146.
[42] SONG Y T, ZHOU D W, LI Q, et al. Leaf nitrogen and phosphorus stoichiometry in 80 herbaceous plant species of Songnen grassland in northeast China [J]. Chin J Plant Ecol, 2012, 36(3): 222–230. doi: 10. 3724/SP.J.1258.2012.00222.宋彦涛, 周道玮, 李强, 等. 松嫩草地80种草本植物叶片氮磷化学计量特征[J]. 植物生态学报, 2012, 36(3): 222–230. doi: 10.3724/ SP.J.1258.2012.00222.
[43] WRIGHT I J, DONG N, MAIRE V, et al. Global climatic drivers of leaf size [J]. Science, 2017, 357(6354): 917–921. doi: 10.1126/science. aal4760.
[44] HU Y S, YAO X Y, LIU Y H. Specific leaf area and its influencing factors of forests at different succession stages in Changbai Mountains [J]. Acta Ecol Sin, 2015, 35(5): 1480–1487. doi: 10.5846/stxb201310 132459.胡耀升, 么旭阳, 刘艳红. 长白山森林不同演替阶段比叶面积及其影响因子[J]. 生态学报, 2015, 35(5): 1480–1487. doi: 10.5846/stxb 201310132459.
[45] POORTER H, NIINEMETS Ü, POORTER L, et al. Causes and conse- quences of variation in leaf mass per area (LMA): A meta-analysis [J]. New Phytol, 2009, 182(3): 565–588. doi: 10.1111/j.1469-8137.2009. 02830.x.
[46] DING J, WU Q, YAN H, et al. Effects of topographic variations and soil characteristics on plant functional traits in a subtropical evergreen broad-leaved forest [J]. Biodiv Sci, 2011, 19(2): 158–167. doi: 10. 3724/SP.J.1003.2011.10312.丁佳, 吴茜, 闫慧, 等. 地形和土壤特性对亚热带常绿阔叶林内植物功能性状的影响 [J]. 生物多样性, 2011, 19(2): 158–167. doi: 10. 3724/SP.J.1003.2011.10312.
Response of Leaf Functional Traits of Shrubs to Altitude inCommunities in Mt. Jinggangshan, Jiangxi, China
XIANG Lin1, CHEN Fang-qing1*, GENG Meng-ya1, WANG Yu-bing1, LÜ Kun1, YANG Shi-lin2
(1. Hubei International Scientific and Technological Cooperation Center of Ecological Conservation and Management in Three Gorges Area, China Three Gorges University,Yichang 443002, Hubei, China; 2. Hubei Zhengjiang Environmental Science & Technology Co., Ltd, Yichang 443002, Hubei, China)
In order to understand the adaptive mechanism ofcommunity in Jinggang Mountain, Jiangxi Province, the changes in leaf functional traits of dominant species in shrub layer were analyzed along altitude. The results showed that the altitudinal gradient had significant effects on leaf functional traits of shrub species. The dry matter content (LDMC), thickness (LT), nitrogen (LNC) and phosphorus concentration (LPC) of leaves increased with altitude rising, whereas the specific leaf area (SLA) and N/P reduced, and the leaf size (LS) increased at first and then decreased. There were significant relations among leaf functional traits. LDMC had negative relation with SLA and LS, positive relation with LT, LNC and LPC. SLA and LT, LNC and LPC had positive relation, whereas LS and LT, N/P and LPC had negative relations, respectively. Environment factors influence on leaf functional traits. Except for altitude, slope also affected SLA, LPC and N/P, and aspect and gradient influenced LS and LNC, respectively. So, it was suggested that the shrub species incommunity could adapt altitude and other environment factors by adjusting leaf functional traits.
Leaf; Functional trait; Altitude; Dry matter;; Jinggang Mountain
10.11926/jtsb.3930
2018-04-18
2018-07-02
国家科技基础性工作专项(2015FY110300)资助
This work was supported by the Special Project for Basic Works of National Science and Technology (Grant No. 2015FY110300).
向琳(1993~ ),女,硕士研究生,研究方向为恢复生态学。E-mail: 1351258603@qq.com
E-mail: fqchen@ctgu.edu.cn
