大气CO2浓度升高和干旱的交互作用对大豆光合作用的影响
2019-03-14李炳言郝兴宇
李炳言,王 娜,郝兴宇,李 萍
(山西农业大学农学院,山西太谷 030801)
工业革命以来人类大量使用石化燃料,导致全球大气CO2浓度持续升高[1]。目前,大气中CO2浓度已由工业革命前的280 μmol/mol增加到406 μmol/mol,并且还以每年0.5%的速度增长,预计21世纪末,可增加到 421~936 μmol/mol[2]。CO2等温室气体增加将导致全球气温上升,降水格局也将发生变化,未来我国大范围干旱等极端天气发生的频次和强度将增加[3]。根据2013年政府间气候变化专门委员会第5次评估报告(IPCC 5,2013):由陆地和海洋表面温度变化的线性趋势得出,全球年平均气温增幅为0.85℃[4],而且最近50 a内我国的平均气温增幅为1.1℃,比全球同期的增加速率明显提高。水资源短缺是目前制约农业生产的一个全球性问题,全球约43%的耕地处于干旱、半干旱地区,我国干旱半干旱地区占全国土地面积的52.5%,其中,半干旱地区占21.7%[5]。干旱是我国西北和东北的西部地区农业发展最大的限制因子。
CO2是绿色植物进行光合作用获取能量的重要原料之一。CO2浓度升高一方面会提高植物的净光合速率,减少蒸腾作用,另一方面会抑制植物的光呼吸过程,增加植物在水分胁迫条件下的抗旱性,有利于光合产物的积累,促进生长[6]。大豆在我国是仅次于水稻、小麦、玉米的经济作物[7],是豆科植物中对干旱较为敏感的一种作物[8],在其整个生育期内需水量较高。严重的干旱条件会使大豆的蛋白质、油量等品质指标下降,造成大豆减产。研究表明,大豆初花期前,适度干旱对其影响不大,在开花期至结荚鼓粒期,过度干旱则会导致植株矮小,籽粒品质下降,产量出现一个较大幅度的降低[9]。目前,对CO2浓度升高与干旱互作的研究较少,其机制有待深入探讨[10]。
本研究通过开展CO2浓度升高和干旱互作条件下大豆光合生理变化的研究,旨在为未来气候变化背景下大豆生产提供理论依据。
1 材料和方法
1.1 试验材料
供试大豆品种为中黄35,具有高产、生育期短、抗性强等特点,由中国农业科学院作物科学研究所培育。
1.2 试验设计
试验在山西晋中市太谷县(37.42°N,112.58°E)山西农业大学试验基地进行。该基地具备2个独立的开顶式气室,用于控制环境中CO2浓度,分别是:对照气室(CK,外界大气CO2浓度,设定为400μmol/mol)和处理气室(T,外界大气 CO2浓度+200 μmol/mol,设定为 600 μmol/mol)。土壤水分作为副处理,设2个水平,分别是:干旱(D,35%~45%的相对土壤含水量,为干旱胁迫)和湿润(W,60%~80%的相对土壤含水量,此为适宜水量)。
土壤为褐潮土,装箱前对土壤过筛,除去大颗粒固体杂物。2014年6月16日大豆种植于60cm×40 cm×35 cm的塑料整理箱中,箱的底部打4~5个孔用于排水通气,箱内土壤高度为25 cm,每箱种植大豆10穴,每穴播2~3粒种子,出苗后对其进行间苗处理,每穴留苗1株,设置6次重复。每个气室中干旱和湿润处理各种植8盆大豆。每箱中施入11.04 g N和12.24 g P。全生育期内每隔2 d在8:00—9:00 用便携式土壤水分测定仪(SU-LB,中国)测定土壤含水量,较低时对其进行适量的灌溉处理。
1.3 测定项目及方法
开花期(播后35 d),使用美国Li-6400便携式光合气体分析系统测定大豆叶片光合参数[11],测定时间为9:00—11:00。使用红蓝光源测定,光强设置为1 400 μmol/(m2·s),叶温28℃。随机从每箱中选取有代表性植株1株,共8株,叶片选择倒数第1片完全展开的功能叶,测定净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr),计算水分利用效率(WUE)。

播后36 d,于9:00—14:00每个处理选取3片最上层完全展开的功能叶用Li-6400测定A-Ci曲线,光强设置为1 400 μmol/(m2·s),叶温28℃,设定CO2浓度梯度分别为 400,300,200,100,50,400,600,700,800,1 000,400 μmol/mol。
叶绿素荧光参数的测定:开花期(播后35 d),随机选取倒数第1片完全展开的功能叶[12],用脉冲制式荧光仪(PAM2000,德国)测定和计算叶绿素荧光参数。在8:30—11:30的入射光子通量密度(PPED)下,测量了光适应状态的最大荧光产额(Fm')和最小荧光产额(F0')。在同一天 23:00—1:00,研究了暗适应状态下的最大荧光产率(Fm)和最小荧光产率(F0)。用于测量饱和荧光的高光闪光灯的PPED为4 000 μmol/(m2·s),持续时间为800 ms。测定指标:叶绿素荧光的其他参数,包括PSⅡ光化学的最大光量子效率(Fv/Fm)、PSⅡ的有效光量子效率(Fv'/Fm')、光化学淬灭系数(qP)、非光化学淬灭系数(NPQ)、电子传递速率(ETR)和PSII光化学有效量子产率(ΦPSⅡ)。
1.4 数据处理
试验数据的处理、图表的绘制用Excel整理,通过SPSS软件中的多元方差分析法进行显著性分析。
2 结果与分析
2.1 CO2浓度升高与干旱互作对大豆叶片光合作用的影响
2.1.1 CO2浓度升高与干旱互作对大豆叶片净光合速率的影响 从图1可以看出,干旱使开花期大豆叶片净光合速率下降。CO2浓度升高使大豆叶片净光合速率较CK增加,湿润条件下增加了40.3%,干旱条件下增加3.3%,干旱条件下增幅明显小于湿润条件下增幅。
2.1.2 CO2浓度升高与干旱互作对大豆叶片气孔导度的影响 从图2可以看出,干旱使开花期大豆气孔导度下降。CO2浓度升高对开花期大豆气孔导度影响不显著,但干旱条件大豆的气孔导度有降低的趋势。

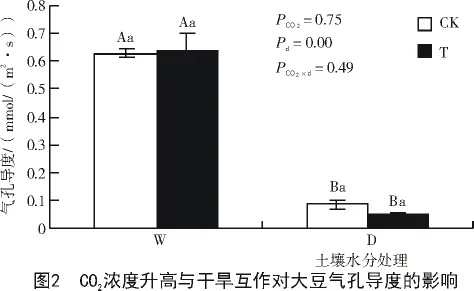
2.1.3 CO2浓度升高与干旱互作对大豆叶片蒸腾速率的影响
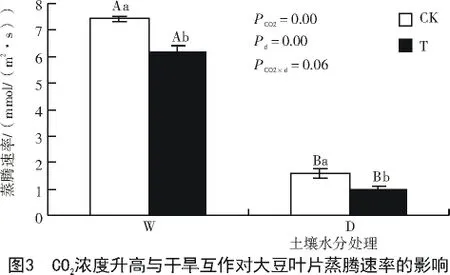
由图3可知,干旱条件会极显著降低大豆的蒸腾速率。CO2浓度升高使湿润和干旱条件下蒸腾速率分别较CK显著下降17.1%和37.5%,二者间无明显互作效应。
2.1.4 CO2浓度升高与干旱互作对大豆水分利用效率的影响 干旱条件会显著提高大豆的水分利用效率。CO2浓度升高使湿润和干旱条件下水分利用效率分别较CK显著提高了71.3%和47.4%。高CO2浓度与干旱有明显互作效应,湿润条件下水分利用效率增幅较干旱条件下增幅更高(图4)。

2.2 CO2浓度升高与干旱互作对大豆叶绿素荧光参数的影响

表1 CO2浓度升高与干旱互作条件下大豆的叶绿素荧光参数变化
从表1可以看出,干旱条件会极显著降低大豆的 Fv'/Fm',ΦPSⅡ,ETR,qP,显著提高 NPQ。CO2浓度升高对大豆叶绿素荧光参数影响不显著。干旱和CO2浓度升高无显著互作效应。
2.3 开花期大豆的A-Ci曲线
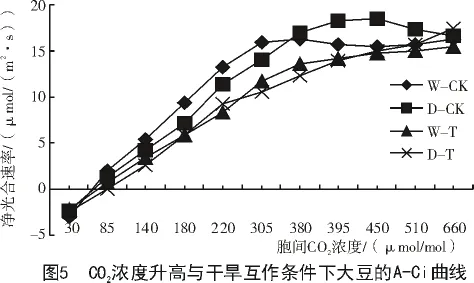
由图5可知,随着胞间CO2浓度的增大,大豆的净光合速率增加。胞间CO2浓度在380 μmol/mol以下湿润条件的净光合速率均高于干旱条件下净光合速率(相同胞间CO2浓度下)。相同胞间CO2浓度条件下,处理气室(高CO2浓度)大豆净光合速率小于对照气室(正常CO2浓度)。
3 结论与讨论
光合作用为绿色植物所特有,植物的生长发育依赖光合作用中形成的营养物质。植物的光合作用极大的受限于外界的环境条件[13]。通过影响净光合速率和呼吸速率等调控植物的生长、发育,进而影响产量及其品质性状。
大气CO2浓度升高后,在湿润条件下大豆的净光合速率显著增加,而在干旱条件下增幅小于湿润条件。可能的原因是:一方面,目前大气中的CO2含量相对较低,尚未达到大豆植物的CO2饱和点,CO2浓度成为大豆光合作用的主要限制因子[14]。因此,在充足的水分供应条件下,提高CO2浓度,有助于增加大豆的净光合速率。另一方面,干旱条件下,大豆叶片的气孔关闭,减小了叶肉细胞内的CO2浓度,而CO2是大豆(C3植物)光合作用极其重要的原料之一。CO2浓度在胞内降低,势必会减弱CO2与O2竞争核酮糖-1,5-二磷酸羧化酶-加氧酶的能力,从而导致大豆光呼吸增强,光合作用减弱[15]。这也是本试验在大气CO2浓度升高和干旱互作中,干旱条件下增加净光合速率较少的原因。
气孔是大豆与外界环境进行气体交换的通道,外界CO2通过气孔进入叶肉细胞,进行光合作用。而气孔导度表示气孔张开的程度,影响光合作用和蒸腾作用。前人研究结果表明,温度会显著影响气孔导度,温度较高不利于植物光合产物的积累,气孔导度降低,反之,气孔导度升高,同时气孔导度也与光照强度有关[16]。本研究发现,开花期大豆叶片气孔导度未受大气CO2浓度影响,但与土壤水分关系密切。干旱条件下,大豆叶片气孔导度极显著降低。CO2浓度升高未降低大豆叶片气孔导度,但会促进大豆叶片进行光合作用,有利于光合产物的积累。而前人研究发现,低浓度CO2会促进气孔张开,高浓度CO2使气孔关闭[17]。造成研究结果差异的原因可能是:(1)CO2进入气孔的途径中相关因子水平受限;(2)CO2浓度和O2浓度在胞内的比值与光合作用中酶的结合位点有关;(3)外界光照影响;(4)CO2进入叶片受到的环境阻力差异。产生这一现象的根本原因有待进一步分析和研究。
干旱和大气中高CO2浓度都会显著降低大豆叶片的蒸腾速率,提高水分利用率。干旱条件下,植物体内的渗透调节物质脯氨酸、甜菜碱等含量增加,叶片蒸腾速率的降低,有助于植物抵抗不良环境[18]。大气CO2浓度升高虽然提高了开花期大豆叶片光合速率和水分利用效率,但干旱条件下增幅小于湿润条件下,表明CO2浓度升高对大豆抗旱能力的提升效果有限。
叶绿素荧光动力学技术由于其能较好的反映光系统对光能的吸收、传递、耗散等参数,通常被测定用于反映植物光合能力的变化。Fv/Fm表示植物对光能的转换效率,Fv'/Fm'表示原初光能捕获效率。ΦPSII反映PSⅡ激发能占总的激发能的比例,荧光淬灭包括光化学淬灭(qP)和非光化学淬灭(NPQ),分别表示PSⅡ中心的开放程度和植物体的热能耗散[19]。本研究中发现,大气CO2浓度升高未显著改变大豆PSⅡ开放中心中的Fv/Fm,Fv'/Fm',ΦPSⅡ,ETR,qP,或是仅表现微弱的增加,NPQ有减小的趋势。而高CO2浓度会显著提高大豆的净光合速率,净光合速率的增加可能与大豆固氮作用有关[20];干旱显著降低 Fv'/Fm',ΦPSⅡ,ETR 和 qP,增加NPQ。这与大豆在干旱胁迫条件下,光合能力的下调相一致,逆境条件下大豆会表现出自身的保护机制,减少光能的转化,减少PSⅡ激发态能量的产生,增加热能的耗散过程;大气高CO2浓度和干旱没有互作效应,表明开花期大气CO2浓度升高对大豆干旱缓解效应有限。
A-Ci曲线表明,相同胞间CO2浓度条件下,高CO2浓度条件下大豆净光合速率小于对照气室叶片净光合速率,这可能与长期高CO2浓度升高导致的光适应有关[21]。
总之,大气CO2浓度会增加开花期大豆光合速率和水分利用效率,在湿润条件下增幅更高。未来的大气CO2浓度升高对开花期大豆抗旱能力提升效果不明显。
