AM真菌的异核现象对共生系统的影响研究进展
2019-03-14王艳胜王爱萍郝艳平梁月秀赵立松尹梦娇张春来
王艳胜 ,耿 帅 ,王爱萍 ,郝艳平 ,梁月秀 ,赵立松 ,尹梦娇 ,张 腾 ,温 贺 ,张春来
(1.山西农业大学农学院,山西省旱作农业与作物生态重点实验室,山西太谷 030801;2.山西农业大学林学院,山西太谷 030801)
AM真菌即从枝菌根真菌[1],通过其孢子产生的菌丝、丛枝以及泡囊与主要的陆生植物形成共生关系[2],真菌通过菌丝从宿主植物中获得碳源,植物从菌根中吸收丰富的微量矿质元素[3],特别是植物不易吸收到的磷、锌、铜等微量元素。研究结果表明,AM真菌对多种经济作物具有增产效果,可提高植物的品质,如增加生物量和植物有效成分含量[4];增强植物的抗逆性,如改善植物的水分状况[5]、提高植物的抗旱能力[6]、提高植物抵抗低温的能力、耐盐碱胁迫的能力[7];增强宿主植物对重金属离子的耐性;还有降低其他真菌、线虫、细菌等病害的作用[8]。目前对共生体系中植物的遗传特性了解较多[9],但由于AM真菌是专性寄生生物,目前尚未获得纯培养,对AM真菌的遗传特性知之甚少。AM真菌是多核的,数百个胞核共存于胞内,尽管有争议,但直接或间接的证据都表明,在单个的AM真菌中包含有大量的遗传性不同的核[10-12]。此外,在同一个核中已观察到遗传差异,这就说明了AM真菌个体内部高度的遗传变异性[12]。近来有证据表明,AM真菌有异形核[13]。
为了解异形核对AM真菌的遗传特性和对共生系统的影响,笔者综述了有关与共生关系建立和共生功能相关的基因,分析了AM真菌异核性可能的原因及其对共生系统即真菌和植物的生态和进化的影响,并展望了AM真菌遗传特性的研究前景。
1 与共生建立和共生功能相关的基因
AM真菌是最常见的一种内生菌根真菌,在与植物的有机结合体系中,真菌菌丝首先附着在植物根表面上,然后侵入植物根表皮细胞(epidermal cells),最后到达皮层细胞(cortical cells)[14]。通过形成丛枝结构(arbuscules)和膨大的泡囊结构(vesicles)在植物皮层细胞中定殖,与其相连的根外菌丝和孢子则分布在根表面或根际土壤中,这样菌丝体及其各种结构就形成了传输根外营养物质到根内的桥梁,并扩大了植物根的吸收表面积[14]。丛枝结构是AM真菌与宿主植物营养物质交换的场所,植物从菌根中吸收丰富的微量矿质元素,而真菌的碳源则从宿主植物中获取[3]。另外,真菌菌丝可同时侵入一种或多种不同植物,使得在一个共生系统中通过形成地下菌丝网连接着许多不同植物。菌丝末端也可形成孢子,这些孢子萌发可形成新的共生体。
一些与菌根共生关系形成的关键阶段和共生关系作用的相关基因的鉴定已经取得了很大进展。共生关系形成大体分为共生前期、侵染初期、延伸到根皮层、形成丛枝结构、共生关系建立共5个阶段[15]。迄今,已鉴定的共生关系作用的相关基因均来源于植物,而非真菌。在共生前期,真菌菌丝附着于根,可观察到许多菌丝分枝。有人认为,从类胡萝卜通路中合成的植物激素——独角金内酯(Strigolactones),是共生的化学信号,可诱导菌丝分枝,引起真菌线粒体功能异常[16-17],但是与之相关的基因目前并不清楚。真菌产生的识别信号物质——Myc因子,诱导植物基因ENOD11表达,ENOD11在蒺藜状苜蓿(Medicago truncatula)中编码富含脯氨酸的假想的多重细胞壁蛋白,该基因在菌根形成的早期和后期,甚至是已形成丛枝的细胞中都有表达[18]。最近研究发现,为了引导真菌到达植物皮层细胞,植物体细胞内构建了一个通道,称之为预侵染器,真菌通过它延伸到植物细胞,植物细胞核随着通道的发育迁移[19-20]。研究表明,蒺藜状苜蓿中的2个基因DMI2和DMI3对诱发预侵染器是必需的。利用豆科模式植物百脉根(Lotus japonicus)突变体已经确定,CASTOR,POLLUX,NUP85,NUP133,CYCLOPS 等5种基因对真菌在豆科植物体内的发育和丛枝的发育是必需的。这些基因在根瘤菌-豆科植物共生体系和菌根共生体系中都是常见的[15]。研究发现,这些基因在水稻(Oryza sativa)的菌根形成中也是必需的。在水稻中鉴定了CASTOR,POLLUX,CCAMK(DMI3),CYCLOPS的直系同源基因,发现这些基因对水稻和豆科植物的菌根形成都是必需的[21]。然而,也发现了另一些基因的作用仅限于水稻菌根形成。它们可分为2组:一组在共生关系发展早期丛枝形成前被表达,有AM1-3和AM11;另一组在发育后期和形成丛枝的细胞中表达,有AM10-11,AM14-15,AM18,AM20,AM24,AM25,AM26,AM29,AM31,AM34,AM39,AM42 和 PT11,这些基因编码的功能目前还不清楚。
目前,AM真菌中的2个基因可能与真菌在植物根内的发育有关。因为没有适合真菌的基因敲除或转化方法,最成功的方法是在AM真菌中寻找直系同源基因,这些基因对植物根被真菌病原体成功地侵染很重要。研究表明,STE12的直系同源基因和类GTP酶可能在根侵染和随后菌丝在根内生长的调控分别起作用[22-23]。因为丛枝是植物和真菌相互作用的桥梁,是营养交换的场所,对含丛枝结构的细胞上基因的表达尤为关注。研究发现,菌根根系提取物中的亲脂性信号物质能够诱导马铃薯磷酸转运基因StPT3和 StPT4的表达[24],而这2个基因是被AM真菌专性诱导表达的。这就说明溶血磷脂作为信号物质在植物与AM真菌共生中起着特别重要的作用。这些磷酸盐转运蛋白在丛枝中表达,因而对共生功能起作用。已被证实,含丛枝结构的细胞中存在一些其他的植物磷酸盐转运蛋白和一种真菌磷酸盐转运蛋白。真菌中其他一些对共生系统发育和共生功能比较重要的基因也已被证实。AM真菌中超氧化物歧化酶基因GmarCuZnSOD表达可以增强植物对氧化物胁迫的耐受性。此外,在根外菌丝中磷转运蛋白也参与了将磷酸盐转运到植物中[25]。
2 AMF异核性可能的原因
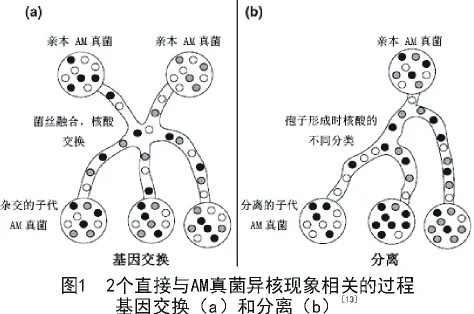
近来有证据表明,AM真菌有异形核,即在所含基因上,不同的核共存于同一菌体内,或同一菌丝细胞内或同一孢子内。2个直接与真菌异核现象形成相关的过程被称之为分离和基因交换[13]。
2.1 基因交换
同一孢子的不同菌丝间可融合并创建新的细胞质连接[26],通过这种融合在单个孢子个体中甚至是不同寄主的根之间就建立起一个大的菌丝网[27]。最近有研究表明,在Rhizophagus irregularis中不同孢子的菌丝也可融合[28],融合后2个AM真菌孢子的核酸在菌丝中混合,那么菌丝融合后产生的子代就会继承双亲的遗传物质[28]。而且一些子代的孢子和菌丝密度等一些遗传特性与其亲本和其他子代都不同[28]。AM真菌个体间核酸交换形成了其独特的表型。这种2个AM真菌亲本间核酸的交换并导致AM真菌子代拥有双亲核酸的现象称之为基因交换(图1-a)。融合后,交换了核酸的AM真菌个体被称为“AM真菌亲本”。
基因交换直接导致了AM真菌的异核性。基因交换的一个潜在结果是一个种群中遗传性不同的个体可相互连接并形成一个巨大的遗传性不同的菌丝网。而且遗传性不同的AM真菌个体间核酸的交换对AM真菌个体异质性的维持有重要影响[11,29]。基因交换另一个重要影响是通过基因交换,不同AMF亲本杂合的核酸在子代中增加了核型的遗传多样性。
基因交换可能对AM真菌在一个变化的环境中快速进化的能力有重要影响,不仅通过在选择时提供新的等位基因,而且通过潜在的允许来源存在差异的核酸重组[30]。在AM真菌中当融合事件可能比生殖周期发生的更频繁时,基因交换和分离可能在快速适应异常的环境条件中成为一种重要的方式[31]。这样的水平基因转移一直被认为在细菌中能促进基因组的可塑性和适应性[32]。
2.2 分离
在AM真菌中另一个导致异核现象的原因是一个亲本孢子在孢子形成时由于核型分离出现了拥有不同核酸物质的子代。这种在孢子形成时出现不同的核酸的现象称之为分离(图1-b)。通过分离产生的后代称为分离系。AM真菌分离现象的一个潜在结果就是亲本孢子在产生新的子代时由于稀有核型的缺失导致AMF遗传多样性降低。基因分离直到不久前都认为是不可能发生的,以前的研究通过观察G.etunicatum中共存的一个基因PLS的13种变异型的遗传研究[33],发现PLS基因的13种变异型存在于所有的子一代中,研究者通过子一代一个位点变异的分离研究得出,在AM真菌中没有发生分离,然而多个位点经过多代生殖后是否发生分离未可知。不久前在R.irregularis中发现分离现象[10],并指出分离发生的可能性依赖于亲本核型的遗传多样性。通过隔离和分别培养来自一个亲本基因交换所得的子一代的单个孢子来研究分离过程的试验已经完成。分离减少了一个新形成种群中等位基因个体大小,从而有助于不同的等位基因的相对丰度的遗传漂变,促进表型异常[8],融合可能有助于等位基因改组,促进异核现象的维持。
3 AM真菌的异核现象对共生系统的影响
已经在AM真菌物种水平上深入研究了AM真菌多样性对植物多样性和植物生产力的影响以及不同寄主对AM真菌孢子形成和不同AM真菌群体共存的影响[34]。然而,AM真菌的异核现象对AM真菌遗传多样性的影响及其对真菌与植物相互作用的影响则很少被关注。通过基因交换和分离使得子代相关核型频率的改变在菌丝网中经常出现。基因交换和分离对共生有着非常重要的影响。最近有文献研究了遗传交换和分离对植株生长和基因表达的影响[10,35]。与之前的转化根的体外培养相比,这些研究是在温室中完整的植株个体上完成的,试验对植株分别接种基因交换所得孢子、分离所得孢子以及亲本孢子,经过几个月的温室培养后,测其干质量及植物基因的遗传转录,研究结果均表明,通过分离和遗传交换产生的遗传变异对水稻生长和基因表达都有显著影响。然而,后续研究则表明,通过基因交换和分离获得的子代,与其亲本和其他子代相比,可以有差异地改变植物的生长和植物基因转录[10]。由不同的AM真菌系引起的对植物生长的改变会因为寄主的改变而不固定。研究表明,水稻接种某一些分离所得的孢子比接种亲本孢子或其他一些分离所得孢子增产5倍[10]。这一研究结果也表明,基因交换和分离对水稻基因的表达产生了很大影响,同时发现接种同样的AM真菌孢子,对水稻的生长有促进作用,但并未促进车前草生长。总之,由于异核性,AM真菌对不同植物个体的生长有不同的影响,需要研究者们进一步探索。
异核性使得AM真菌有快速产生可变后代的能力,与亲本相比,增加了其产生不同适应性子代的可能性,从而对新的环境条件适应的能力。如此灵活的基因型和表型可能是有性生殖快速的可替代的选择,也可能是AM真菌生态学成功的关键。
AM真菌的基因型随着环境的变化而改变,环境决定AM真菌的基因型。物种间的相互作用和适应性反馈被认为是在自然界中允许植物和AM真菌物种多样性共存的机制。研究表明,这一机制也可用于解释比AM真菌低等的物种,并可在一个AM真菌物种中发现基因型和表型多样性。甚至,这一机制可作用于AM真菌个体规模,参与AM真菌内部个体基因多样性的维持。基因交换与分离对孢子的核型含量有重要作用。这些机制在很短的时间内就可以导致子代异常的共生效果。一些新的AM真菌后代由于它们独有的特异核型,可能比其他的AM真菌更适合改变了的环境,因此,被选择下来。
异核现象可以通过在很短的时间内创造各种各样的新的AM真菌子代并筛选出来,这样可以增强AM真菌的适应性。环境的异质性,如不同的寄主种类,可以影响基因交换和分离形成的不同核型的AM真菌的命运,可以潜在的有助于AM真菌中异核现象的维持。
4 展望
AM真菌是最丰富的植物共生体,可提高植物生产力和多样性。遗传物质交换可能对AM真菌在一个变化的环境中快速进化的能力有重要影响,基因交换和重组可能在快速适应异常的环境条件中成为一种重要的方式。接下来的挑战将是去调查自然界中AM真菌种群融合事件发生的频率,以及它们是如何通过不同的AM真菌世代广泛分布的。虽然一些与自然条件有更多相关的原位系统研究已取得重要进展[36-37],但还缺乏微生态相关的环境来估计融合频率。另一个研究方向是去调查是否有来自AM真菌的重组是由于隐藏着的有性生殖活动,多个减数分裂基因和假定的性信息素已经在Glomus spp 被证实了[38-39]。
在AM真菌个体中许多基因组共存的个体和它们传递的下一代,对基因组进化论的理解有深远的影响。那么,基因交换和分离是如何影响真菌在植物根内的分布和发育的?一个真菌的不同基因组是如何作用于他们的生态学、进化论和共生效率的?这些问题有待进一步研究来回答。
