干旱胁迫对云南松苗木生长及碳酸酐酶的影响
2019-03-11王晓丽徐志鸿韦文长和润喜陈诗曹子林
王晓丽,徐志鸿,韦文长,和润喜,陈诗,曹子林
干旱胁迫对云南松苗木生长及碳酸酐酶的影响
王晓丽1,徐志鸿1,韦文长2,和润喜1,陈诗1,曹子林3*
1. 西南林业大学林学院, 云南 昆明 650224 2. 双江县林业局, 云南 临沧 677399 3. 西南林业大学生态与水土保持学院, 云南 昆明 650224
以云南松苗木为研究材料,采用称重法控制土壤含水量对苗木进行干旱胁迫,探讨不同干旱胁迫强度对云南松苗木生长形态指标、生物量及碳酸酐酶活性的影响。结果表明:(1)相对于针叶长来说,地径对干旱胁迫更为敏感,程度较小的干旱胁迫即可引起地径生长的显著降低;随着干旱胁迫程度的加大,主根长呈先增加后减小的变化趋势,中度干旱胁迫(T4)是主根长生长的转折点。(2)高强度干旱胁迫对苗木生物量的抑制效应在器官间表现不均衡,苗木根受到的抑制作用更强;中度干旱胁迫(T4)处理时苗木根生物量比最大,同时叶生物量比最小,一定程度的干旱胁迫有利于苗木对根和叶生物量比的调整。(3)随着干旱胁迫强度的增大,苗木根、茎、叶的碳酸酐酶活性皆呈逐渐下降的趋势,方差分析和多重比较表明,可根据根中的碳酸酐酶活性来初步筛选耐旱性更强的优良单株。总之,该年龄段苗木的地径越大、主根长越长、根生物量比越大、根部的碳酸酐酶活性越高,苗木的耐旱能力越强,越有利于苗木在石漠化生境中的成活和生长。
云南松; 苗木形态指标; 生物量; 碳酸酐酶; 干旱胁迫
石漠化(Stony Desertification)是中国西南岩溶地区脆弱岩溶基底基础上形成的一种特有荒漠化生态现象。云南地处中国西南石漠化中心地区,是中国岩溶分布最广的省区之一。云南岩溶发育,往往导致土壤表层干旱缺水、土壤浅薄贫瘠(缺磷少氮)、岩石裸露、地形破碎、生态环境脆弱[1-3],已成为云南岩溶地区最大的生态环境问题,是制约该区经济、社会与生态可持续发展的障碍。在岩溶区,根据基岩裸露率、坡度、土壤厚度、土被覆盖度、土被加植被盖度等因子,将石漠化划分为无明显石漠化、潜在石漠化、轻度、中度、重度、极度石漠化6个等级[4]。云南松()林作为我国西南地区特有的森林植被类型,是云南省主要森林类型[5],云南松天然林主要分布区的石漠化等级为无明显石漠化、潜在石漠化、轻度石漠化和中度石漠化,可见,云南岩溶地区是云南松天然林分布的主要区域之一,云南松在岩溶生境中具有极强的协同适应性。
在岩溶地区,人们已经认识到植被对岩溶发育具有强大的驱动力,有报道认为碳酸酐酶(Carbonic Anhydrase,简称CA)对碳酸盐岩的溶解具有催化作用。在岩溶动力系统中加入CA可使石灰岩的溶解速率提高10倍[6-11];云南松体内的CA在岩溶生境中具有良好的稳定性以及云南松体内CA活性与林内土壤中CA活性存在极显著的相关性[12,13];硅藻[14]分泌的CA能促进石灰岩的溶蚀,因此,植物体内CA在生物岩溶发育过程中扮演着重要的角色。另外,研究发现CA在光合作用途径的碳固定中起着重要的作用,可加速无机碳向羧化酶活性部位的扩散,增加CO2的固定速率[15];岩溶区玉米[16]叶片的CA活性和光合作用也高于非岩溶区。CA活性高低对植物水分利用及光合作用都有较大影响,干旱胁迫下番茄CA活性变化对植株光合作用及水分利用的影响研究[17]表明番茄叶水势和光合速率与CA表达活性之间存在一定的变化趋势相似性。CA可看作植物适应干旱胁迫[17,18]、盐胁迫[19]、低温胁迫[20]等胁迫条件的光合碳代谢调节酶, 在保持光合碳代谢稳定方面具有重要功能。综上所述,CA在植物生物岩溶和逆境适应性方面皆具有重要的作用。生物岩溶和逆境适应性是岩溶地区植被恢复及石漠化治理研究的核心内容,所以可将CA研究作为一个重要的切入点,开展云南岩溶生境条件下植被CA活性及其生物岩溶作用机制的相关研究,为云南岩溶地区植被恢复及石漠化治理提供一种新的理论视角,是解决云南岩溶地区生态系统恢复与重建的重要途径。
目前,以云南岩溶生境的云南松林为研究对象,对云南松CA在岩溶生境中的稳定性和云南松CA活性及其与林内土壤中CA活性的相关性做了一些基础性的前期研究工作[12,13],但是,关于岩溶环境因子对云南松CA活性的影响、岩溶生境下云南松CA活性的变化规律以及与CA活性密切相关的云南松生长指标研究却缺乏,不仅制约人们对云南松岩溶环境适应性及其生物岩溶作用机制的理解,而且不利于云南松天然林可持续发展、石漠化生态恢复等实践活动。本文以7个月生云南松苗木为试验材料,采用称重法控制土壤含水量,研究干旱胁迫(主要岩溶环境因子之一)对云南松苗木形态指标、生物量及CA活性的影响,探讨云南松苗木生长指标及CA活性随主要岩溶环境因子的变化规律,以CA为切入点,分析云南松苗木的岩溶生境适应性和其生物岩溶作用机制。为现有云南松天然林的可持续发展和云南松人工林的定向培育提供理论依据和技术支撑,同时丰富我国岩溶生态系统良性生态恢复的机理研究。
1 材料与方法
1.1 研究材料
以7个月生云南松苗木为试验材料,云南松种子来自于楚雄双柏的云南松天然林,母树年龄30 a左右,生长健壮,干形通直。
1.2 试验地概况
试验地位于云南省昆明市西南林业大学温室内的苗圃地。昆明处于中国西南部,云贵高原中部,位于东经102°10′~103°40′,北纬24°23′~26°22′,东西最大横距140 km,南北最大纵距220 km;地势为北高南低,平均海拔约1894 m,年平均气温14.7 ℃左右,绝对最低气温-9 ℃,绝对最高气温32.5 ℃,年降雨量700~1100 mm,年平均相对湿度68.2%,属中亚热带气候类型[21],为云南松的天然分布区。
1.3 研究方法
1.3.1 苗木培育2016年8月,在温室中进行容器育苗,育苗容器为单体硬质聚乙烯盆状容器,内径为22 cm,高度为20 cm,每盆装2 kg土壤,土壤为取自云南松林下的山地黄壤,每盆播50粒经过催芽处理的云南松种子,播后覆土,浇水。待苗龄3个月时进行间苗并定苗,每盆留生长一致的苗木30株,期间进行正常的水分管理。
1.3.2 苗木干旱处理2016年11月,采用完全随机试验设计,将已定苗的48盆苗木随机分成6组,每组8盆,设置充分供水(T1)(对照)、正常水分(T2)(对照)、轻度干旱胁迫(T3)、中度干旱胁迫(T4)、重度干旱胁迫(T5)和极重度干旱胁迫(T6)6个水分处理梯度,每个处理8盆苗木;每五天,采用称重法补充自然流失的水分,使土壤含水量保持在各个胁迫范围内。T1、T2、T3、T4、T5和T6处理的土壤水分含量依次为田间最大持水量的90%~95%、75%~80%、60%~65%、45%~50%、30%~35%和15%~20%[22,23]。对供试苗木持续做4个月的干旱胁迫处理。
1.3.3 试验材料取样方法2017年3月,将各干旱胁迫处理的所有的云南松苗木全部从盆中取出,尽可能保持植株及其根系的完整,尽快将根部清洗干净并去除干枯的针叶,然后将植株表面的浮水擦干并放入装有冰块的保温箱中进行暂时冷藏,以备后续带回实验室进行苗木形态指标、生物量和碳酸酐酶活性测定使用。
1.3.4 苗木形态指标和生物量的测定每个处理三个重复,每个处理取30株苗木,用直尺测定其主根长、苗高、针叶长(每株随机测定三根针叶,取平均值),用游标卡尺测定其地径,数出一级侧根数,上述形态指标测定完成后,将各处理植株的根、茎、叶分开并分器官称重,然后分别计算根、茎、叶的生物量比。根生物量比=(根鲜重/苗木总鲜重),茎生物量比=(茎鲜重/苗木总鲜重),叶生物量比=(叶鲜重/苗木总鲜重)。
1.3.5 碳酸酐酶提取各处理分别取云南松新鲜根、茎、叶各7 g,剪碎放入预冷的研钵中,加入液氮,再加入21 mL巴比妥提取缓冲液(10 mmol/L,含巯基乙醇50 mmol/L,pH8.3)进行研磨,取研磨液倒入10 mL离心管中,离心管置于冰浴中20 min后,在4 ℃条件下,5000 r/min离心10 min,取上清液即为粗提液,冷藏待测[12,13,24],每个处理每个器官三个重复。
1.3.6 碳酸酐酶活性测定云南松苗木各器官碳酸酐酶活性的测定采用pH计法[12,13,24,25]。保持反应系统在0~2 ℃,取待测粗提液1 mL,加入到含15 mL的巴比妥缓冲液(20 mmol/L,pH8.3)的反应容器中,然后迅速加入10 mL预冷的(0~2 ℃)饱和CO2蒸馏水,用pH电极监测反应体系pH值的变化,记下pH下降1个单位(8.2至7.2)所需时间,记为t,同时记录同一样品在酶失活条件下pH下降1个单位所需的时间,记为0,酶的活性用-unit表示[26,27]。=(t/0-1)。酶相对活性:(活性/对照活性)×100%。
1.3.7 数据分析运用SPSS软件进行方差分析和多重比较(Duncan氏新复极差法)[26-28]。
2 结果与分析
2.1 干旱胁迫对云南松苗木生长形态指标的影响
随着水分处理梯度的变化,云南松的苗高、针叶长两个指标的变化趋势基本一致,即随着水分的减少,苗高和针叶长皆表现先增加后减小,虽然正常水分(T2)(对照)处理是水分丰缺的转折点,但是轻度干旱胁迫(T3)更有利于苗高和针叶的生长。地径的变化情况与苗高、针叶长有所不同,随着水分的减少,地径变小,中度干旱胁迫(T4)处理地径达最小值。方差分析表明,地径存在显著差异,针叶长存在极显著差异;多重比较显示,中度干旱胁迫是地径显著降低的转折点,重度干旱胁迫是针叶长显著降低的转折点,相对与针叶长来说,地径对干旱胁迫更为敏感,程度较小的干旱胁迫即可引起地径生长的显著降低(表1)。
干旱胁迫下的云南松苗木的主根长高于正常水分(T2)(对照)处理的主根长,一定程度的干旱胁迫会促进主根往土壤深层的生长;中度干旱胁迫(T4)处理的主根长极显著的大于正常水分(T2)(对照)和极重度干旱胁迫(T6)处理的主根长,在此干旱处理下苗木从增加主根长来调节其抗旱能力已达其上限,如果干旱胁迫程度进一步加深,则主根长降低。同时,一级侧根数量在轻度干旱胁迫(T3)时达到峰值,该水分条件更有利于侧根的产生和生长,而后随着干旱胁迫程度的加深,一级侧根数量降低(表1)。
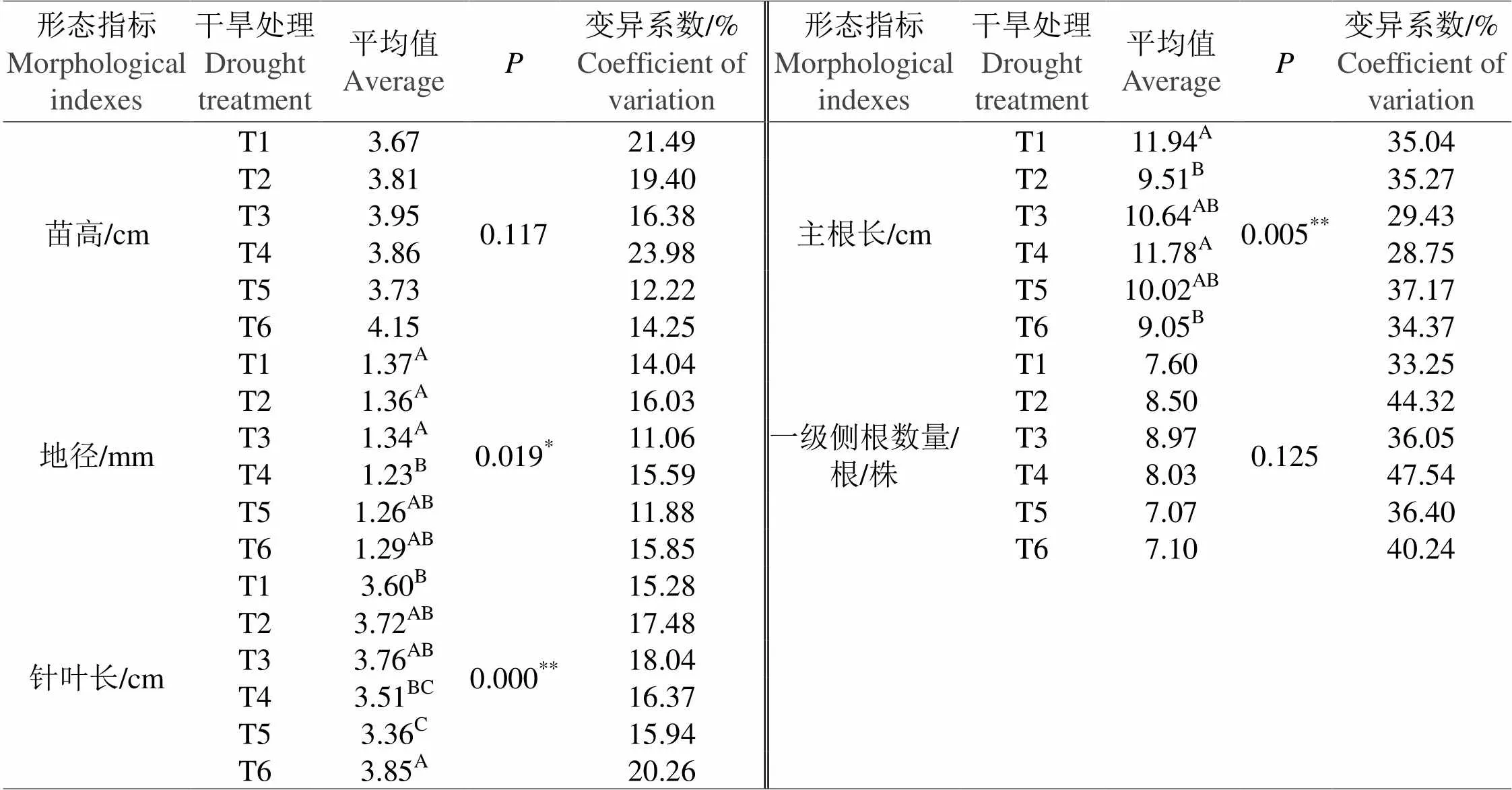
表 1 不同干旱胁迫处理间云南松苗木生长形态指标的方差分析和多重比较
备注:*代表处理间差异显著;**代表处理间差异极显著;数字右上角的字母代表两两处理间的差异显著情况。
Note:*shows significant difference between the representatives; ** shows extremely significant difference between the representatives; the letters in the upper right corner represent the difference between the two treatments.
2.2 干旱胁迫对云南松苗木生物量的影响
2.2.1 干旱胁迫对云南松苗木生物量鲜重的影响云南松苗木根生物量鲜重的变化趋势为:随着干旱胁迫程度的加深,生物量鲜重呈现下降的情况,且皆低于对照处理;云南松苗木茎、叶及总生物量鲜重的变化趋势基本相同,即随着干旱胁迫程度的加深,生物量鲜重总体上皆呈先平缓下降后升高的趋势(图1)。随着大强度干旱胁迫的作用,抑制效应在器官间表现不均衡,大强度的干旱胁迫对根生长的抑制影响大于其对茎和叶的影响。
2.2.2 干旱胁迫对云南松苗木生物量分配的影响在不同干旱胁迫处理下,云南松苗木根和茎的生物量所占总生物量的比例大体相当,都在20%左右,叶的生物量所占比例较高,在60%左右,虽然该比例不会随着干旱胁迫强度的增大出现很大的波动,但是在干旱处理中,中度干旱胁迫(T4)处理时根生物量比最大,相应的叶生物量比最小(图2),苗木对根和叶生物量比的调整更有利于其抗逆能力的提高,更高的干旱胁迫强度不利于苗木从生物量分配调整方面增强其抗逆能力,这与干旱胁迫对云南松苗木主根长的分析结果一致。

2.3 干旱胁迫对云南松苗木碳酸酐酶活性的影响
在干旱胁迫下,云南松苗木根、茎、叶的碳酸酐酶活性的变化规律基本一致,即随着干旱胁迫强度的增大,碳酸酐酶活性皆呈逐渐下降的趋势,且在一定干旱胁迫范围内的相同干旱胁迫处理强度下,叶的碳酸酐酶活性较根、茎的下降幅度更大(表2)。云南松苗木根、茎、叶中的碳酸酐酶活性在不同干旱胁迫处理间皆存在极显著差异,根、叶中碳酸酐酶活性处理间的多重比较皆表明不同干旱胁迫处理间的酶活性没有显著差异,但重度干旱胁迫(T5)和极重度干旱胁迫(T6)处理的根中的酶活性与对照(T1、T2)处理的酶活性间皆具有极显著差异,且中度干旱胁迫(T4)、重度干旱胁迫(T5)和极重度干旱胁迫(T6)处理的叶中的酶活性与对照(T1)处理的酶活性间皆存在极显著差异;茎中碳酸酐酶活性处理间的多重比较说明极重度干旱胁迫(T6)处理的酶活性与对照(T1、T2)处理的酶活性间皆存在极显著差异,所有的干旱胁迫处理间的酶活性不存在显著差异。综上所述,从云南松苗木不同器官的碳酸酐酶活性角度分析,根中的酶活性可以作为云南松苗木耐旱性筛选的主要指标,因此可根据根中的碳酸酐酶活性来初步筛选耐旱性更强的更适应岩溶生境的优良单株。

表2 不同干旱胁迫处理间云南松苗木碳酸酐酶活性的方差分析和多重比较
备注:*代表处理间差异显著;**代表处理间差异极显著;数字右上角的字母代表两两处理间的差异显著情况。
Note:*shows significant difference between the representatives; **shows extremely significant difference between the representatives; the letters in the upper right corner represent the difference between the two treatments.
3 讨论
干旱胁迫下,云南松苗木各形态指标和生物量指标总体皆呈下降的趋势,此变化情况与前人在文冠果[23]、白刺花[22]、构树[18]、桑树[18]等树种上所做的干旱胁迫效应方面的研究结果基本一致。虽然CA在植物生物岩溶和逆境适应性方面皆具有重要的作用,但前人对CA的研究主要集中在CA在光合作用途径中的作用研究[15],认为CA是光合碳代谢调节酶, 在保持光合碳代谢稳定方面具有重要功能,从而增强植物的逆境适应性。然而对于植物通过生物岩溶作用来改善其立地条件和增强其逆境适应性方面的研究很少,本文作者以云南岩溶生境的云南松林为研究对象,对云南松CA在岩溶生境中的稳定性和云南松CA活性及其与林内土壤中CA活性的相关性做了一些基础性的前期研究工作,发现云南松体内的CA在岩溶生境中具有良好的稳定性以及云南松体内CA活性与林内土壤中CA活性存在极显著的相关性[12,13],因此云南松体内CA的活性与云南松的生物岩溶能力密切相关。将前期研究与本文研究相结合,认为在干旱胁迫(主要岩溶生境因子之一)下,可根据云南松苗木中(主要是根中)的碳酸酐酶活性来初步筛选耐旱性更强的更适应岩溶生境的云南松优良单株。
受研究时间和试验材料的限制,本文仅以7个月生的云南松苗木为研究材料,由于干旱处理时间整体较短,因此在不同器官生物量分配研究方面有一定的局限性(由于苗木较小,生物量较少,各器官的生物量更少,所以本研究采用了鲜重指标,而没有采用干重指标。),研究结论只代表了该阶段的不同器官生物量分配策略,后续随着苗木的生长和林木的发育,其不同生长阶段在干旱胁迫中的生物量分配策略还有待进一步的深入研究,才能比较全面的认识云南松的干旱胁迫的生物量分配策略。另外,干旱胁迫虽然是岩溶生境的主要环境因子,但岩溶生境同时还存在土壤浅薄贫瘠(缺磷少氮)的问题,因此后续可开展土壤理化性质对云南松生长和碳酸酐酶活性影响方面的研究,综合分析岩溶生境中云南松的适生策略,为进一步筛选适应不同等级石漠化区域的优良云南松单株、家系或无性系提供理论依据和技术支撑。
4 结论
对不同干旱胁迫处理下地上部分形态指标的分析表明地径对干旱胁迫更为敏感,相对与针叶长来说,程度较小的干旱胁迫即可引起地径生长的显著降低。对不同干旱胁迫处理下地下部分形态指标的分析表明,中度干旱胁迫(T4)是主根长生长的转折点,在此干旱处理下苗木从增加主根长以增强对土壤水分的吸收来调节其抗旱能力已达其上限。因此在潜在石漠化、轻度石漠化和中度石漠化区域,可以通过苗木地径指标和主根长指标来初步筛选出适生的云南松优良单株。不同干旱胁迫处理对云南松苗木生物量鲜重影响的变化说明,高强度的干旱胁迫对根生长的抑制影响大于其对茎和叶的影响。生物量分配方面,中度干旱胁迫(T4)处理时根生物量比最大,相应的叶生物量比最小,云南松苗木对根和叶生物量比的调整更有利于其抗逆能力的提高。因此生物量指标的研究结果与上述形态指标的研究结果一致。随着干旱胁迫强度的增大,云南松苗木不同器官根、茎、叶的碳酸酐酶活性皆呈逐渐下降的趋势,方差分析和多重比较表明,可根据根中的碳酸酐酶活性来筛选耐旱性更强的更适应岩溶生境的云南松优良单株。综合云南松苗木形态指标、生物量指标和碳酸酐酶指标的研究结果,认为该年龄段的苗木的地径越大、主根长越长、根生物量比越大、根部的碳酸酐酶活性越高,苗木的耐旱能力越强,越有利于苗木在石漠化生境中的成活和生长。
[1] 王宇,张贵.滇东岩溶石山地区石漠化特征及成因[J].地球科学进展,2003,18(6):933-938
[2] 赖兴会.云南石漠化的生态特征及其危机表现[J].林业调查规划,2004,29(2):80-82
[3] 张云,周跃华,常恩福.云南省石漠化问题初探[J].林业经济,2010(5):72-74
[4] 白晓永,熊康宁,苏孝良,等.喀斯特石漠化景观及其土地生态效应——以贵州贞丰县为例[J].中国岩溶,2005,24(4):276-281
[5] 蔡年辉,李根前.云南松天然林区植物群落结构的空间动态研究[J].西北植物学报,2006,26(10):2119-2124
[6] 潘根兴,曹建华.表层带岩溶作用:以土壤为媒介的地球表层生态系统过程—以桂林峰丛洼地岩溶系统为例[J].中 国岩溶,1999,18(4): 287-295
[7] Song LH, Liang FY. Distribution of CO2in soil air affected by vegetationin the shilin national park[J]. Acta Geologica Sinica, 2000,75(3):288-293
[8] Tripp BC, Smith K, Ferry JG. Carbonic anhydrase: new insights for an ancient enzyme[J]. J Biol Chem, 2001,276:48615-48618
[9] Cronk JD, Endrizzi JA, Cronk MR,. Crystal structure of E. coli alpha-carbonic anhydrase, an enzyme with an unusual PH-dependent activity[J]. Protein Sci, 2001,10:911-922
[10] Lind SS. Structure and mechanism of carbonic anhydrase[J]. Pharmacol Ther, 1997,74:1-20
[11] Kimber MS, Pai EF.The active site architecture of Pisumsativum beta-carbonic anhydrase is a mirror image of that of alpha-carbonic anhydrase[J]. EMBO J, 2000,19:1407-1418
[12] 王晓丽,曹子林,和润喜,等.不同pH、温度和储藏因素对云南松针叶碳酸酐酶稳定性的影响[J].云南大学学报:自然 科学版,2016,38(3):501-506
[13] 王晓丽,曹子林,和润喜,等.云南松根叶及林内土壤的碳酸酐酶活性分析[J].西南林业大学学报,2016,36(2):31-34
[14] 王倩,支崇远.硅藻碳酸酐酶对石灰岩岩溶的作用及其生态意义[J].上海地质,2007(4):25-27
[15] 蒋春云,马秀灵,沈晓艳,等.植物碳酸酐酶的研究进展[J].植物生理学报,2013,49(6):545-550
[16] 胥献宇.不同温度条件下玉米碳酸酐酶活性差异比较与分析[J].种子,2010,29(3):84-88
[17] 孙卫红,吴秋霞,温新宇,等.干旱胁迫下番茄叶片碳酸酐酶活性的变化[J].植物生理学报,2015,51(4):424-428
[18] 吴沿友,梁铮,刑德科.模拟干旱胁迫下构树和桑树的生理特征比较[J].广西植物,2011,31(1):92-96
[19] 何宣,王白羽,张晓磊,等.盐胁迫下小黑麦碳酸酐酶基因表达及其酶活性分析[J].新疆农业科学,2012,49(7):1197-1202
[20] 陈虎,何新华,罗聪,等.低温胁迫下龙眼碳酸酐酶基因(CA)的克隆与表达分析[J].园艺学报,2012,39(2):243-252
[21] 黄红春,杨志杰,文斌,等.2016年昆明城市绿化植物冻害调查及恢复研究[J].园林科技,2016,142(4):15-22
[22] 李芳兰,包维楷,吴宁.白刺花幼苗对不同强度干旱胁迫的形态与生理响应[J].生态学报,2009,29(10):5406-5416
[23] 谢志玉,张文辉,刘新成.干旱胁迫对文冠果幼苗生长和生理生化特征的影响[J].西北植物学报,2010,30(5):948-954
[24] 吴沿友,李西腾,郝建朝,等.不同植物的碳酸酐酶活力差异研究[J].广西植物,2006,26(4):366-369
[25] 曾宪东,余龙江,李为,等.西南岩溶地区黄荆叶片碳酸酐酶的稳定性[J].植物学通报,2005,22(2):169-174
[26] 刘明磊.正交试验设计中的方差分析[D].哈尔滨:东北林业大学,2011
[27] 罗传义,时景荣,戴传波.基于Excel的正交试验方差分析程序[J].计算机工程,2002,28(11):240-24
[28] 徐仲安,王天保,李常英,等.正交试验设计法简介[J].科技情报开发与经济,2002,12(5):148-150
Effect of Drought Stress on Growth and Carbonic Anhydrase Activity ofSeedlings
WANG Xiao-li1, XU Zhi-hong1, WEI Wen-chang2, HE Run-xi1, CHEN Shi1, CAO Zi-lin3*
1.650224,2.677399,3.650224,
This study examined the effects of drought stresses on the morphological indexes and carbonic anhydrase activity ofseedlings. The results were as following: 1) The ground diameter was the most sensitive to drought stress compared with the needle length of the seedlings. The low level of drought stress can result in a significant decrease in ground diameter increment. The taproot length increased to maximum at medium treatment and then decrease along the levels of drought stresses. 2) The changes in roots, stems, leaves and total biomasses inseedlings generally decreased along the levels of drought stresses. The inhibiting effect of drought stress on root growth was greater than that on stems and leaves. The effect of drought stress on biomass proportion was ranked as leaves > stems ≈ roots. The roots were more sensitive to the increasing drought stress intensity with the highest adjustment of root ratio in the seedlings observed at the treatment of moderate drought stress. Whereas, higher levels of drought stresses were not conducive to the improvement of their resilience in terms of the biomass allocation. 3) The activities of carbonic anhydride in roots, stems and leaves ofseedlings decreased with the increase of intensity drought stress. The results from variance analysis and multiple comparisons showed that the activity of carbonic anhydrase in roots was an important prediction for the drought tolerance. We concluded that longer taproot length, and higher ground diameter, root biomass, and carbonic anhydrase activity of roots may be important implication for the evaluation of drought-resistant ability, as well as the capacity of survival and growth ofseedlings in rocky desertification habitat.
; seedling morphological indexes; biomass; carbonic anhydrase; drought stress
Q948
A
1000-2324(2019)01-0006-06
10.3969/j.issn.1000-2324.2019.01.002
2017-06-27
2017-08-07
云南省高校优势特色重点学科(生态学)建设项目(515006083);“十三五”国家重点研发计划课题项目(2016YFD0600501)
王晓丽(1976-),女,在读博士研究生,副教授,主要从事森林培育和林木生理生态方面的教学与研究工作. E-mail:1144607944@qq.com
Author for correspondence. E-mail:fjcaozilin@qq.com
