2016-2018年广州市H6亚型禽流感病毒外环境分离株HA基因遗传特征分析
2019-02-28李魁彪鲁恩洁陈艺韵刘艳慧
曹 蓝,李魁彪,鲁恩洁,陈艺韵,刘艳慧,夏 丹,狄 飚
禽流感病毒属于正粘病毒科流感病毒属A型流感病毒,根据病毒表面抗原血凝素(HA)和神经氨酸酶(NA)的不同,分为不同亚型:H1~H16,N1~N9[1]。H6亚型禽流感病毒属于低致病性禽流感病毒,禽类感染后常表现为轻微临床症状或无临床症状。1965年首次从火鸡体内分离到H6亚型禽流感病毒以来,该亚型病毒不断地在野禽和家禽体内分离到。研究显示,我国早期H6亚型禽流感病毒主要为H6N1和H6N2[2],近期主要流行亚型为H6N6和H6N2[3],同时其流行和传播呈上升趋势,是我国南方地区家禽常见的禽流感病毒亚型之一[4]。一方面H6亚型禽流感病毒可以为人感染H5亚型禽流感病毒提供基因骨架,进而增加禽流感病毒跨物种传播风险[5],另一方面H6亚型禽流感病毒可以直接感人,具有突破种间屏障而感染哺乳动物甚至感染人的潜能[6]。当前我国家禽中H6与H5、H7、H9亚型共流行,增加了不同禽流感病毒重组的复杂性。研究发现,HA蛋白受体结合位点变异在禽流感病毒跨宿主传播中发挥了重要作用[6]。本研究以广州地区2016-2018年外环境监测分离的H6亚型禽流感病毒为研究对象,分析HA基因变异和进化特点,进而监测广州地区H6亚型禽流感病毒跨物种传播风险。
1 材料与方法
1.1样本来源 根据广州市外环境禽流感病毒监测方案,在禽类生鲜市场、活禽肉菜市场、批发市场、家禽规模养殖场(户)或散养户集中地区和野生禽鸟栖息地等五类场所中,随机选取粪便、饮水、污水、笼具、刀具、地面、称、脱毛机等不同类型外环境标本并于4 ℃保存,在24 h内送至实验室于-80 ℃冰箱保存备用。2016-2018年的监测标本中有6份标本经荧光定量RT-PCR检测为A型、H6亚型禽流感病毒阳性,H5、H7和H9亚型禽流感病毒阴性,检测试剂盒购于江苏硕世生物科技股份有限公司。
1.2病毒分离 阳性标本进行10 000 g离心2 min后取上清,抗生素处理后接种9~10 d龄SPF鸡胚。37 ℃培养2~3 d后,收获接胚尿囊液,通过血凝实验记录病毒滴度,血凝阳性毒株再一次经荧光定量RT-PCR检测为A型、H6亚型禽流感病毒阳性,H5、H7和H9亚型禽流感病毒阴性。
1.3序列测定 应用Oligo 6软件设计H6亚型禽流感病毒HA基因全长扩增引物,引物由华大基因合成。通过RT-PCR方法扩增HA基因,将阳性鉴定产物送至广州华大基因公司,通过ABI 3730进行病毒基因一代测序。
1.4序列分析 以2013年台湾分离的人源毒株A/Taiwan/2/2013(H6N1)和近年H6亚型病毒流行株HA基因作为比较基因,应用DNA Star7.1软件拼接HA基因序列并对序列进行同源性分析,用MegAlign软件比对受体结合位点和糖基化位点变异情况。使用MEGA 4.0软件,以HA基因ORF(Open Reading Frame)为基本单元,从GISAID(Global Initiative on Sharing All Influenza Data)数据库下载相关参考序列,绘制HA基因系统进化树。绘制方法为Neighbor-joining法(参数设置为1 000 replications)及Maximum composite likelihood model比对核苷酸序列。
2 结 果
2.1同源性分析 6株病毒HA基因核苷酸同源性在81.0%~99.1%之间,分离株HA蛋白氨基酸同源性在81.3%~99.4%之间,分离株核苷酸最大相似性毒株见表1。
表1 2016-2018年广州H6亚型禽流感病毒HA基因最大相似性毒株
Tab.1 Viruses of maximum similarity with HA gene of H6 subtype avian influenza viruses in Guangzhou from 2016 to 2018

毒株名称最大相似性毒株相似度/%A/environment/2016B1395/2016A/duck/Jiangxi/262/2013(mixed)97A/environment/2017NX25557/2017A/duck/Ganzhou/GZ151/2016(H6N6)99A/environment/2018XN03891/2018A/duck/Ganzhou/GZ151/2016(H6N6)98A/environment/20180467/2018A/duck/Ganzhou/GZ151/2016(H6N6)98A/environment/20180518/2018A/duck/Ganzhou/GZ151/2016(H6N6)98A/environment/2018XN13565/2018A/pigeon/Guangxi/158/2014(H6N2)97
2.2分子特征分析 有研究表明,HA裂解位点附近具有4个以上连续的碱性氨基酸插入是高致病性禽流感病毒重要的分子标志[7],本研究的6株毒株的裂解位点位于第345位和第346位氨基酸之间,且裂解位点序列相对保守,没有出现变异,只在第345位有1个碱性氨基酸,呈现为低致病性禽流感病毒分子特点。HA蛋白第226位和228位是禽流感病毒重要的受体结合位点,本研究中6株毒株均为226Q和228G,即倾向与禽类呼吸道细胞受体结合,此外A/environment/2018XN13565/2018毒株出现E190T变异。H6亚型禽流感病毒HA蛋白共有8个糖基化位点,其中A/environment/2016B1395/2016毒株在183位出现糖基化位点缺失,详见表2。
表2 2016-2018年广州H6亚型禽流感病毒HA基因分子特征
Tab.2 Molecular characteristics of HA gene of H6 subtype avian influenza viruses in Guangzhou from 2016 to 2018
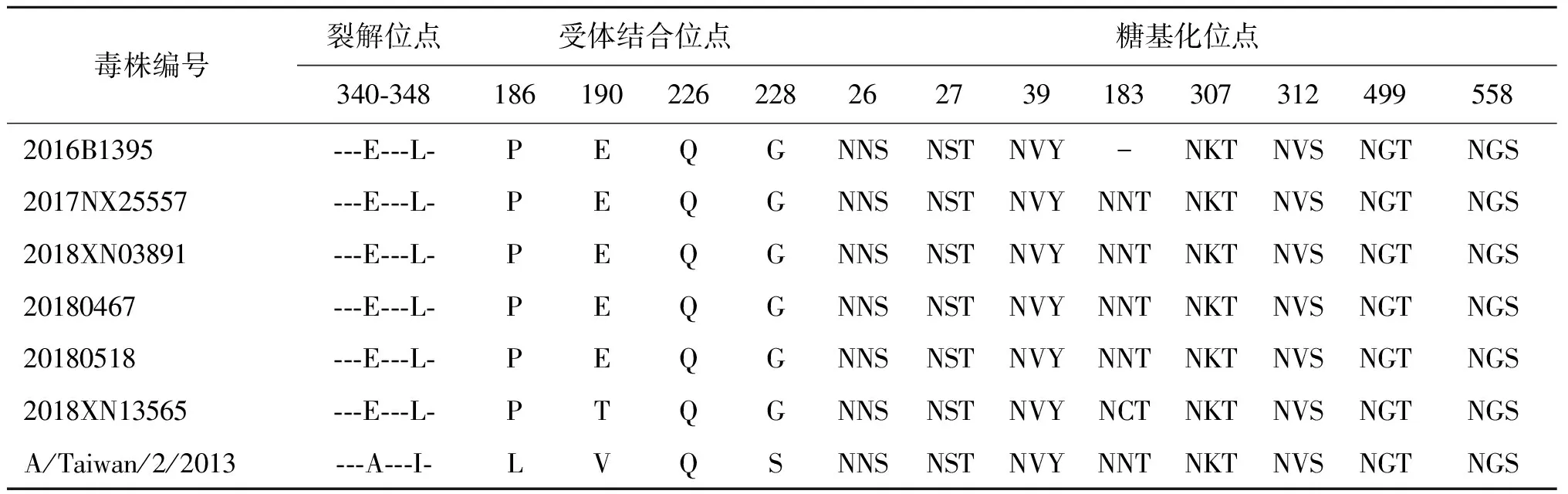
毒株编号裂解位点受体结合位点糖基化位点340-3481861902262282627391833073124995582016B1395---E---L-PEQGNNSNSTNVY-NKT NVSNGTNGS2017NX25557---E---L-PEQGNNSNSTNVYNNTNKT NVSNGTNGS2018XN03891---E---L-PEQGNNSNSTNVYNNTNKT NVSNGTNGS20180467---E---L-PEQGNNSNSTNVYNNTNKT NVSNGTNGS20180518---E---L-PEQGNNSNSTNVYNNTNKT NVSNGTNGS2018XN13565---E---L-PTQGNNSNSTNVYNCTNKT NVSNGTNGSA/Taiwan/2/2013---A---I- LVQSNNSNSTNVYNNTNKT NVSNGTNGS
2.3遗传进化分析 本研究中2016-2018年5株毒株归属于欧亚谱系内的ST2853-like分支,该分支内流行毒株分离地的不同,又形成2个小分支,其中A/environment/2016B1395/2016毒株与华东地区流行毒株亲缘关系相近,其余4株毒株与华南地区流行毒株亲缘关系相近。而2018年另有一毒株(A/environment/2018XN13565/2018)归属于欧亚谱系内的ST339-like分支,见图1。
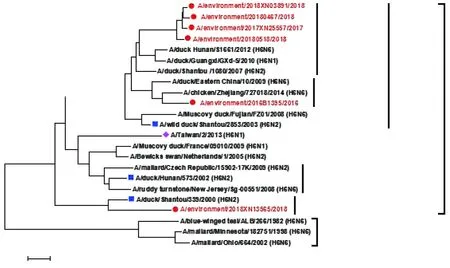
注:“●”代表2016-2018年广州外环境分离株;“■”代表不同进化分支代表株;“◆”代表人源毒株图1 2016-2018年广州H6亚型禽流感病毒HA基因遗传进化分析Fig.1 Genetic evolution analysis of HA gene of H6 subtype avian influenza viruses in Guangzhou from 2016 to 2018
3 讨 论
1997年出现首例人感染H5N1禽流感病例,研究发现H6N1的7个基因片段与该H5N1病毒高度同源,推测H6N1是H5N1的前体病毒,而同样在2014年我国四川出现的首例人感染H5N6禽流感病例中发现,H5N6病毒NA基因即是与H6N6病毒重组而来[8],提示说明H6亚型禽流感病毒可以通过重组而增加病毒的跨物种传播风险,以及H6亚型禽流感病毒与H5亚型禽流感病毒之间发生自然重组的普遍性,推测H5Nx流行株的 NA亚型与H6Nx的NA亚型流行有关。因此当前流行的H5N6病毒,是否会继续以H6Nx为病毒骨架,进一步重组产生新型H5Nx病毒,需要特别关注。活禽市场是禽流感病毒流行和传播的重要场所,Peng等[9]监测研究显示,2012-2016年我国家禽中H6亚型禽流感病毒的流行呈上升趋势。因此开展H6亚型禽流感病毒流行情况及其NA亚型分布情况监测,对监测新型H5Nx的出现及禽流感病毒跨种间传播具有一定的预警意义。
早在2009年的研究报道显示,H6亚型禽流感病毒可以人工感染小鼠和雪貂[10-11],2011年我国广东分离到猪源H6N6禽流感病毒[12],特别是2013年台湾报道了首例人感染H6N1禽流感病毒病例[13],血清学研究也显示,2007年在广西健康人群中检测到了H6亚型禽流感病毒抗体[14],上述研究均提示H6亚型禽流感病毒具有突破种间屏障直接感染哺乳动物甚至感染人的潜能,这对公共卫生构成严重威胁。进一步研究显示,HA蛋白G228S突变增加了H6N1禽流感病毒与人唾液酸受体的结合能力,这是H6亚型禽流感病毒感染人的重要分子基础[6],此外也与HA蛋白受体结合位点P186L突变有关。本研究中,2016-2018年分离株HA基因与人源A/Taiwan/2/2013(H6N1)毒株同源性差异较大(核苷酸同源性在82.8%~83.4%之间),基因比对显示大部分分离株与广东地区H6N6亚型禽源病毒基因相似度最高。但2016-2018年广州分离株之间同源性差异也变化较大,最大同源性高达99.1%,最小同源性仅为81.0%,说明虽然近年来广州地区H6毒株尚为禽源本地化病毒,也未监测到与2013年台湾人源H6N1病毒基因同源的相似病毒存在,但HA基因已出现较大变异,提示持续开展H6亚型禽流感病毒基因进化监测的必要性。
进化分析上,广州H6亚型禽流感病毒环境分离株与人源A/Taiwan/2/2013(H6N1)位于不同进化分支。2018年之前广州分离株遗传进化上相对单一,均属于ST-2853 like流行分支,与以往报道一致[15-16],但由于禽流感病毒基因变异较快,在ST-2853 like分支内,2016年毒株与华东地区流行毒株亲缘较近,2017-2018年毒株与华南地区流行毒株亲缘较近,出现一定的进化差异。从2018年开始,广州分离株保持在ST2853-like分支继续进化的基础上,又出现ST339-like新流行分支病毒。
基因分子变异上,H6亚型禽流感病毒HA蛋白只有1个碱性氨基酸插入,属于低致病性禽流感病毒,虽对禽类呈现低致病性,但临床症状上难于发现,更有利于病毒的潜在传播,提示未来一段时间H6亚型禽流感病毒仍然是活禽市场常见流行的禽流感病毒亚型之一。本研究毒株中HA蛋白受体结合位点相对保守,尚未出现重要变异,为典型的禽类受体位点特点,推测发生跨种间传播的风险较低。
利益冲突:无
