Rapid Creation of New Photoperiod-/Thermo-Sensitive Genic Male-Sterile Rice Materials by CRISPR/Cas9 System
2019-02-19ShenLanDongGuojunZhangYuHuGuochengZhangQiangHuGuanglianXuBoRenDeyongHuJiangZhuLiGaoZhenyuZhangGuanghengGuoLongbiaoZengDaliQianQian
Shen Lan, Dong Guojun, Zhang Yu, Hu Guocheng, Zhang Qiang, Hu Guanglian, Xu Bo, Ren Deyong, Hu Jiang, Zhu Li, Gao Zhenyu, Zhang Guangheng, Guo Longbiao, Zeng Dali, Qian Qian
Rapid Creation of New Photoperiod-/Thermo-Sensitive Genic Male-Sterile Rice Materials by CRISPR/Cas9 System
Shen Lan#, Dong Guojun#, Zhang Yu, Hu Guocheng, Zhang Qiang, Hu Guanglian, Xu Bo, Ren Deyong, Hu Jiang, Zhu Li, Gao Zhenyu, Zhang Guangheng, Guo Longbiao, Zeng Dali, Qian Qian
(; These authors contributed equally to this study)
Rice is an important food crop in China, and the development of hybrid rice is a crucial way to increase grain yield. The creation of dual-purpose nuclear-sterile lines for two-line hybrid breeding has become vital for commercial rice breeding. We constructed the pC1300-2x35S::Cas9-sgRNAexpression vector for editing themale fertility genein two widely compatible rice varieties, 93-11 and Huazhan, by using the CRISPR/Cas9 system. We obtained the marker-free photoperiod-/thermo- sensitive genic male-sterile (P/TGMS) lines in T1generation. According to the experiments in phytotron with four temperature and photoperiod treatments, we found the temperature is the main factor for restoring the pollen fertility ofmutants in 93-11 and Huazhan, and the photoperiod also has some effects on pollen fertility in two different rice backgrounds. The application of cultivating new male-sterile lines by genome editing system will significantly accelerate the rice breeding process.
Rice (L.) is an important food crop for almost half of the world’s population. Hybrid rice technology increases yields by about 20% under normal irrigated conditions in several countries, contributing significantly to the food security(Peng et al, 1999; Ren et al, 2018). Commercial hybrid rice production uses a three-line hybrid rice system and a two-line hybrid rice system (Cheng et al, 2007; Jiang et al, 2017). Compared with three-line hybrid, the two-line hybrid rice system does not require a maintainer line and restorer line for hybrid seed production(Zhou et al, 2014; Chang et al, 2016). Given the obvious advantages of two-line system, it has been widely used in commercial hybrid rice seed production, and the photoperiod-/thermo-sensitive genic male-sterile (P/TGMS) rice materials are useful germplasm resources for two-line hybrid breeding(Zhou et al, 2012; Zhou et al, 2014; Fan et al, 2016). Therefore, enriching the germplasm resources of sterile lines will remain a key challenge for breeders. With advances in technology, many genes have been successfully identified and used in rice breeding by using the clustered regularly interspaced short palindromic repeats (CRISPR)-associated endonuclease 9 (CRISPR/Cas9) system(Zhou et al, 2016; Tang et al, 2017; Shen et al, 2018; Zhang et al, 2018). In this study, we aimed to create new P/TGMS male-sterile rice materials in different genetic backgrounds by the CRISPR/Cas9 system.
The gene, cloned from Guangzhan 63S, one of the most widely used P/TGMS lines in China, was used for the creation of male-sterile rice mutants. Xu et al (2011) found three natural single nucleotide polymorphism(SNP) mutations ofin Guangzhan 63S. Site 71 in the first exon creates a premature stop codon, site 508 in the intron is delimited in non-coding regions, and site 1469 in the fifth exon is a nonsense mutation. Because P/TGMS lines tend to be selected for their combination ability rather than as restorer lines with restore genes. The good male parent plants 93-11 and Huazhan, crossed with male-sterile lines, have high general combining ability (GCA) and are widely used in three-line and two-line rice hybrids. Owing to their high GCA and broad adaptability, 93-11 and Huazhan were chosen as the background materials. We constructed the pC1300-2×35S::Cas9-sgRNAexpression vector and the target site were consistency and specificity in 93-11 and Huazhan (Fig. 1-A and -B), respectively. Owing to the low efficiency of transgenic technology in mostrice varieties, we only screened three homozygous mutants of 16 transgenic plants of each rice variety in T0generation (Fig. 1-C). These mutations ofin 93-11 and Huazhan resulted in the reading frame shift mutations that would lead to premature transcription termination. In this study, we chose the mutants of code 1 in two rice varieties respectively to produce T1generation.
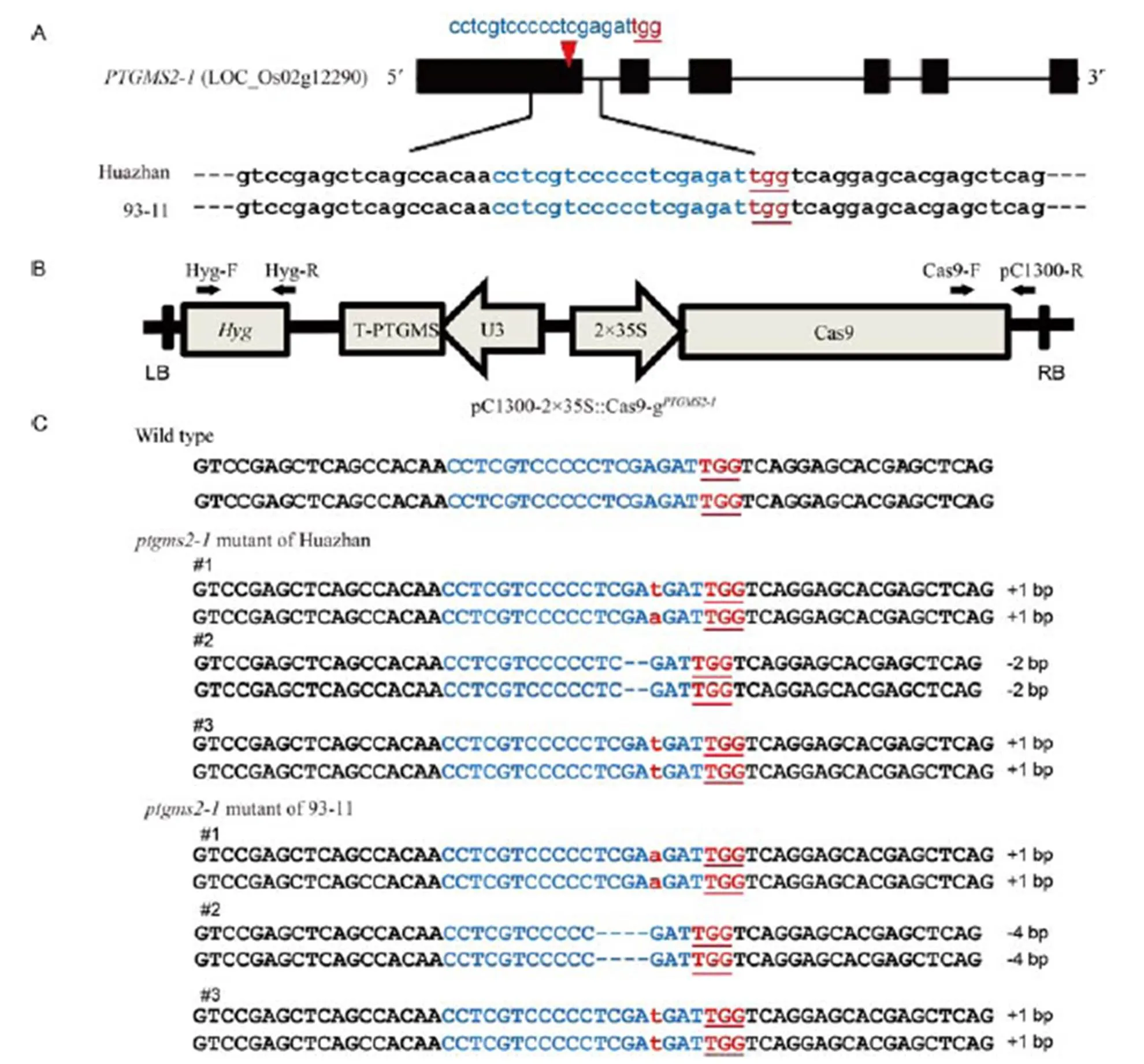
Fig. 1. Schematic diagram of the targeted sites in(A), flow diagram of CRISPR/Cas9 system for editing(B), and mutation types at theloci of Huazhan and 93-11 in the T0generation (C).
The numbers of nucleotide at both sides of the target sequences are 18 bp. Black arrows represent the detection primers.The targeted sequence is highlighted in blue, while the protospacer adjacent motif (PAM) sequence is underlined. Mutations of 1 bp insertions are in red lowercase letters. The deleted sequences are shown by blue hyphens.
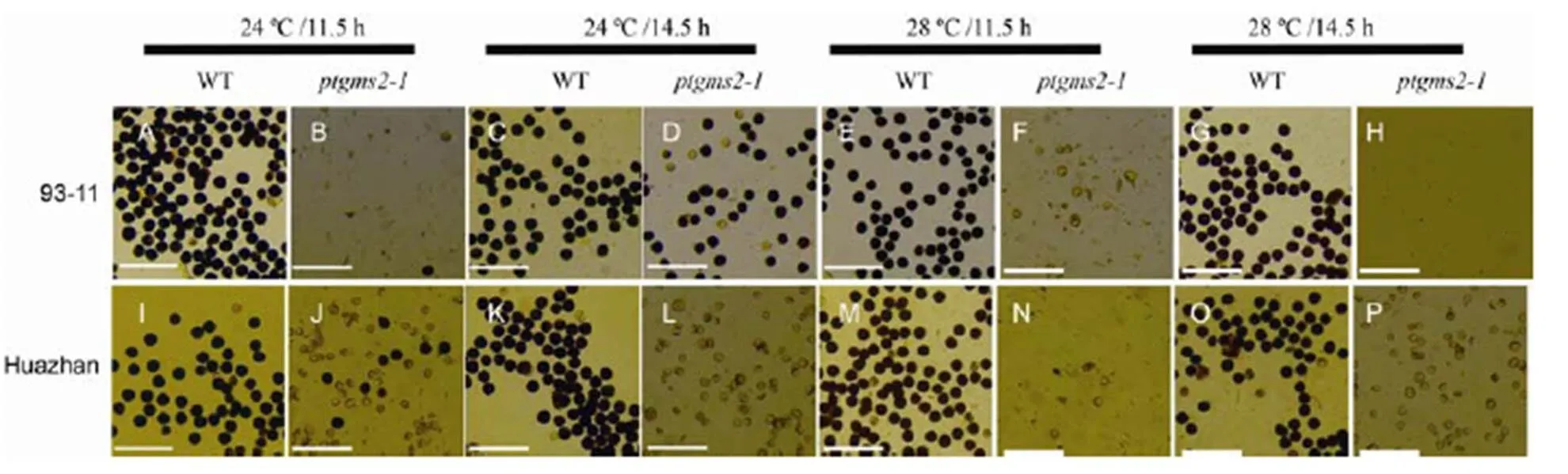
Fig. 2. Pollen fertility of wild type (WT) andin 93-11 and Huazhan.
A–H, I2-KI staining of pollen grains of WT andin 93-11. Scale bars are 200 μm. I–P, I2-KI staining of pollen grains of WT andin Huazhan. Scale bars are 200 μm.
We found thatmutants in 93-11 and Huazhan were almost completely sterile in natural growth, and thesemutants had increased tiller number and significantly decreased plant height than the wild type (WT) (Supplemental Fig. 1-A to -F; Supplemental Fig. 2-A and -B). Next, we found that the panicle length and number of secondary branches per main panicle ofwere significantly decreased in 93-11 (Supplemental Fig. 2-D and -F), while the number of primary branches per main panicle ofwas mildly increased, when compared with WT (Supplemental Fig. 2-E). In Huazhan, the number of secondary branches per main panicle ofwas significantly increased, when compared with WT (Supplemental Fig. 2-J). There was no significant difference in grain number between the WTs and mutants (Supplemental Fig. 2-C and -G). These results suggest that, except for sterility,lines in 93-11 andHuazhan had relatively normal agronomic traits. Simultaneously,we obtained 4 and 3 marker-freemutants of 20 transgenic plants in 93-11 and Huazhan in T1generation (Supplemental Fig. 3). These marker-free mutants in T1generation were chose for further studies.
To determine the critical temperature and photoperiod ofmutants in 93-11 and Huazhan, we used the phytotron to culture plants in T2generation under artificial conditions. The mature florets in wild type of both 93-11 and Huazhan showed normal morphology and fertility under different temperatures and photoperiods (Fig. 2-A, -C, -E, -G, -I, -K, -M and -O; Supplemental Fig. 4). The anthers of all themutants had a distorted shape with a whitish colour compared with the wild type plants (Supplemental Fig. 4). The pollen fertility of the mutants of 93-11 and Huazhan was either decreased greatly, or was even non-existent. Themutant of 93-11 produced no pollen at 28 ºC/14.5 h light (Fig. 2-H), but about 46.7% of pollen grains restored fertility at 24 ºC/14.5 h light (Fig. 2-D). Themutant of Huazhan produced complete abortion pollen at a 14.5 h photoperiod (Fig. 2-L and -P), however, there were a few pollen grains (2.7%, Fig. 2-J), which restored fertility at 24ºC/11.5 h light. We also found that the 11.5 h photoperiod was beneficial for fertility restoration ofmutants of Huazhan at 24 ºC, and under the same low temperature, the 14.5 h photoperiod was more beneficial for fertility restoration ofmutant of 93-11. These results indicate that low temperature is the main factor for restoring the pollen fertility ofmutants of 93-11 and Huazhan, while the photoperiod also has some effects on pollen fertility in two different rice backgrounds. Taken together, we successfully created newlines of 93-11 and Huazhan.
Previous study indicated that the pollen freesterile lines in Guangzhan 63S show temperature-sensitive genetic male sterility (Xu et al, 2011), and other studies showed that mutations ininrice varieties cause thermogenic infertility. Whereas, mutations inrice varieties like Nonken 58S, 7001S and N422S cause photosensitive male sterility (Xu et al, 2011). In this study, we noticed that temperature could be the main factor for the fertility alteration ofin 93-11 and Huazhan under four combinations of photoperiod and temperature. Besides, male fertility mutants of the same gene showed diversity in performance in different rice genetic backgrounds. In our study, the fertility sensitivities ofmutants in the two rice genetic backgrounds to light and temperature are different. The similar results have been found in editinggene of the same target sites in the ZhongzheB (ZZB), ReB, TianfengB (TFB), Wushansimiao (WSSM), Yuejingsimiao (YJSM), Zhenshan 97B (ZS97B) and GAZ backgrounds, which possess different critical sterility-inducing temperature(Zhou et al, 2016). These differences can contribute to the creation of new male-sterile rice materials adapted to the changing environment.
It takes years, even decades, to cultivate new male-sterile lines of diverse rice background under regulated temperature and light conditions in the two-line hybrid rice system using traditional methods (Zhou et al, 2016). However, in our study, marker-free new male-sterile plants could be screened from T1generations (Supplemental Fig. 3). The diversified sterile lines in different genetic backgrounds would enrich the genetic and cytoplasmic diversity. The CRISPR/Cas9 genome editing method to create new P/TGMS plants will explore heterosis more fully and would save considerable time.
In conclusion, we created the new marker-free male fertility mutantsin 93-11 and Huazhan by using the CRISPR/Cas9 system. Our results demonstrate a fast method for the generation of male-sterile plants, and could be used for further hybrid breeding and seed production.
Acknowledgement
This study was supported by the Central Public-Interest Scientific Institution Basal Research Fund of China National Rice Research Institute (Grant No. 2017RG001-4).
SUPPLEMENTAL DATA
The following materials are available in the online version of this article at http://www.sciencedirect.com/science/journal/ 16726308; http://www.ricescience.org.
Supplemental File 1.Materials and methods used in this study.
Supplemental Table 1. Primers used in this research.
Supplemental Fig. 1. Comparison of plant height and tiller number of 93-11 and Huazhan with the mutants in T1generation.
Supplemental Fig. 2. Comparison of panicle-related traits of 93-11 and Huazhan with the mutants in T1generation.
Supplemental Fig. 3. PCR identification of the marker-free transgenic plants in T1generation.
Supplemental Fig. 4. Morphology of the wild type andmutant in 93-11 and Huazhan.
Chang Z Y, Chen Z F, Wang N, Xie G, Lu J W, Yan W, Zhou J L, Tang X Y, Deng X W. 2016. Construction of a male sterility system for hybrid rice breeding and seed production using a nuclear male sterility gene., 113(49): 14145–14150.
Cheng S H, Zhuang J Y, Fan Y Y, Du J H, Cao L Y. 2007. Progress in research and development on hybrid rice: A super-domesticate in China., 100(5): 959–966.
Fan Y R, Yang J Y, Mathioni S M, Yu J S, Shen J Q, Yang X F, Wang L, Zhang Q H, Cai Z X, Xu C G, Li X H, Xiao J H, Meyers B C, Zhang Q F. 2016., producing phased small- interfering RNAs, regulates photoperiod-sensitive male sterility in rice., 113(52): 15144–15149.
Jiang J H, Ni J L, Wu S, Wang D Z. 2017. Development of permanent genic male sterile line by pyramiding thermo-sensitive genic male sterile genes and reverse temperature induced genic male sterile genes in rice (L.)., 31(4): 371–378. (in Chinese with English abstract)
Peng S, Cassman K G, Virmani S S, Sheehy J, Khush G S. 1999. Yield potential trends of tropical rice since the release of IR8 and the challenge of increasing rice yield potential., 39(6): 1552–1559.
Ren D Y, Hu J, Xu Q K, Cui Y J, Zhang Y, Zhou T T, Rao Y C, Xue D W, Zeng D L, Zhang G H, Gao Z Y, Zhu L, Shen L, Chen G, Guo L B, Qian Q. 2018.determines grain size and sterile lemma fate in rice., 69(20): 4853–4866.
Shen L, Wang C, Fu Y P, Wang J J, Liu Q, Zhang X M, Yan C J, QianQ, Wang K J. 2018. QTL editing confers opposing yield performance in different rice varieties., 60(2): 89–93.
Tang L, Mao B G, Li Y K, Lv Q M, Zhang L P, Chen C Y, He H J, Wang W P, Zeng X F, Shao Y, Pan Y L, Hu Y Y, Peng Y, Fu X Q, Li H Q, Xia S T, Zhao B R. 2017. Knockout ofusing the CRISPR/Cas9 system produces low Cd-accumulatingrice without compromising yield., 7(1): 14438.
Xu J J, Wang B H, Wu Y H, Du P N, Wang J, Wang M, Yi C D, Gu M H, Liang G H. 2011. Fine mapping and candidate gene analysis of, the photoperiod-thermo-sensitive genic male sterile gene in rice (L.)., 122(2): 365–372.
Zhang J S, Zhang H, Botella J R, Zhu J K. 2018. Generation of new glutinous rice by CRISPR/Cas9-targeted mutagenesis of thegene in elite rice varieties., 60(5): 369–375.
Zhou H, Liu Q J, Li J, Jiang D G, Zhou L Y, Wu P, Lu S, Li F, Zhu L Y, Liu Z L, Chen L T, Liu Y G, Zhuang C X. 2012. Photoperiod- and thermo-sensitive genic male sterility in rice are caused by a point mutation in a novel noncoding RNA that produces a small RNA., 22(4): 649–660.
Zhou H, Zhou M, Yang Y Z, Li J, Zhu L Y, Jiang D G, Dong J F, Liu Q J, Gu L F, Zhou L Y, Feng M J, Qin P, Hu X C, Song C L, Shi J F, Song X W, Ni E D, Wu X J, Deng Q Y, Liu Z L, Chen M S, Liu Y G, Cao X F, Zhuang C X. 2014. RNase ZS1processes UbL40mRNAs and controls thermosensitive genic male sterility in rice., 5: 4884.
Zhou H, He M, Li J, Chen L, Huang Z F, Zheng S Y, Zhu L Y, Ni E D, Jiang D G, Zhao B R, Zhuang C X. 2016. Development of commercial thermo-sensitive genic male sterile rice accelerates hybrid rice breeding using the CRISPR/Cas9-mediatedediting system., 6: 37395.
(Managing Editor: Wang Caihong)
Copyright © 2019, China National Rice Research Institute. Hosting by Elsevier B V
This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/4.0/)
Peer review under responsibility of China National Rice Research Institute
http://dx.doi.org/10.1016/j.rsci.2018.12.006
30 October 2018;
18 December 2018
Qian Qian (qianqian188@hotmail.com)
杂志排行

Rice Science的其它文章
- Improvements of TKC Technology Accelerate Isolation of Transgene-Free CRISPR/Cas9-Edited Rice Plants
- Production of Two Elite Glutinous Rice Varieties by Editing Wx Gene
- Development and Application of CRISPR/Cas System in Rice
- CRISPR/Cas9-Mediated Adenine Base Editing in Rice Genome
- Targeted Mutagenesis of NAC Transcription Factor Gene, OsNAC041, Leading to Salt Sensitivity in Rice
- Characterization and Evaluation of OsLCT1 and OsNramp5 Mutants Generated Through CRISPR/Cas9-Mediated Mutagenesis for Breeding Low Cd Rice