发芽粟米研究进展
2019-02-15高金燕袁娟丽陈红兵
胡 帅 高金燕 武 涌,3 袁娟丽 陈红兵
(南昌大学食品科学与技术国家重点实验室1,南昌 330047)(南昌大学食品学院2,南昌 330047)(南昌大学中德联合研究院3,南昌 330047)(南昌大学药学院4,南昌 330006)
近年来,随着人们对饮食要求的不断提高及高新食品加工技术的不断发展,发芽作为一种重要的加工技术,因其纯天然,损失小,营养高等特点受到国内外学者的关注。当干燥的谷物吸水溶胀后,休眠的酶被活化导致代谢活性增加,产生初级和次级代谢产物,使谷物发生复杂的物理、化学和结构变化,从而改善谷物的营养和功能特性。发芽能够改变谷物的外观、风味及营养价值。谷物发芽产物可以以芽菜的形式食用或进一步加工后食用,如蒸煮、炒制等,也可以配料添加的方式使用,可以提高产品营养价值。
我国是粟类作物(粟米)生产大国,拥有丰富的粟类资源。2016年粟米产量为两百万吨,位居世界第三,其中谷子占世界总产量的80%以上[1],而且以谷子为主的粟类作物曾是我国餐桌上的主粮。粟芽作为粟米发芽后的产物越来越受到青睐。然而,与国外相比,国内对粟芽在食品中的应用研究相对较少,很大程度上是受制于传统思想束缚及法规的限制。粟芽在我国为一种传统药材,但并未在药食同源中批准使用,导致对粟芽食品研发较为滞后。我国对糙米及小麦等谷物发芽研究与应用已经起步,但粟米发芽研究鲜有涉及,对粟米资源的开发与利用极为不利。本文详细介绍发芽粟米的操作条件、加工性质、营养特性及其在食品中的应用,可为发芽粟米的产业发展提供参考。
1 粟米与粟芽的概述
1.1 粟米与粟芽的定义
关于粟米的定义存在较多的争议,目前主要有广义和狭义两种。广义的粟米是指粟类作物,粟类作物不是植物学分类术语,而是一些籽粒较小的禾谷类作物和草本植物的总称。常见的主要有珍珠粟(Pearl millet,Pennisetumglaucum)、谷子(Foxtail millet,Setariaitalic)、龙爪稷(Finger millet,Eleusinecoracana)、糜子(Proso millet,Panicummiliaceum)、圆果雀稗(Kodo millet,Paspalumscrobiculatum)、稗子(Barnyard millet,Echinochloafrumentaceae)和细柄黍(Little millet,Panicumsumatrense)等。其中珍珠粟产量最高,约占世界粟类总产量的46%[2]。粟米脱壳后为较小的米粒,称为小米,英文millets涵盖了粟米与小米的含义,若要明确区分粟米与小米,则小米可称为dehusked millets。而狭义的粟米则特指谷子(Foxtail millet,Setariaitalic),谷子只是粟类作物的一小部分,小米则为谷子脱壳后的米粒,谷子发芽后即为粟芽(SetariaeFructusGerminatus),是我国一种传统的中药材。《本草纲目》中记载粟芽具有消积食、开胃、除烦、除热功效。《中国药典》中则称粟芽具有消食和中,健脾开胃的功能,主要用于治疗食积不消,腹胀口臭,脾胃虚弱,不饥食少。本文就广义的发芽粟米研究进行相关阐述。
1.2 粟米发芽条件工艺的优化
粟米发芽操作与其他谷物基本相同,主要过程为粟粒筛选清洗→灭菌消毒→浸泡→沥干→发芽→干燥保存,筛选清洗的目的是为了除去不完整颗粒及粟粒表面的污染物,而灭菌消毒一般采用1%次氯酸钠溶液或1.5%甲醛溶液,粟米在水中浸泡沥干后,在人工气候箱或其他受控条件下发芽,最后通过热空气干燥终止发芽。为了提高粟芽的品质,使发芽朝着最优的方向进行,常对发芽条件进行优化(表1、表2),浸泡时间、发芽温度与发芽时间是常见的三个发芽参数。浸泡不仅是种子吸胀的过程,还可以引起种子中一些抗营养成分及矿物质含量的变化,对代谢过程有很大影响。Sihag[3]将珍珠粟在25 ℃下浸泡0~12 h,发现其抗氧化成份、β-胡萝卜素、铁元素及灰分都随着浸泡的时间延长而下降。Bello等[4]在室温下将珍珠粟浸泡24~72 h后,发现其植酸及多酚物质随着浸泡时间而减少。发芽时间和温度范围不仅决定了粟芽中酶活性及代谢活性,还决定了色素和风味化合物的形成。如Hiremath等[5]将龙爪稷发芽24~72 h,结果表明植酸与铁的含量都随时间减少。Obadina等[6]将珍珠粟发芽12~96 h,结果发现其蛋白质和纤维随时间有所提高,而酚类、脂肪和碳水化合物都有所下降。实际上,据文献报道,糜子[7]发芽过程中也有相似的规律。此外,发芽时间的延长和温度的升高会使珍珠粟[8]和龙爪稷[9]中淀粉酶,脂肪酶的活性增强,但在最适时间和温度后发生下降。浸泡后种子的水分含量有时会替代浸泡时间来研究条件对种子发芽的影响,如Swami等[10]探究龙爪稷的吸水动力学时发现,浸泡3 h之后龙爪稷吸水达到饱和,温度能够使浸泡时间缩短,并提高吸水率,在30、40、50、60 ℃的饱和吸水率分别为45.72%、51.05%、56.39%和60.34%,符合Peleg吸水模型[11]。另外,即使是同一品种粟米,不同性状的发芽结果也不一样。如Shukla等[12]发现棕色谷子发芽后淀粉酶活性高于白色品种,而发芽后紫色谷子的脂肪降低率(59.77%)大于黄色谷子(53.49%)[13]。
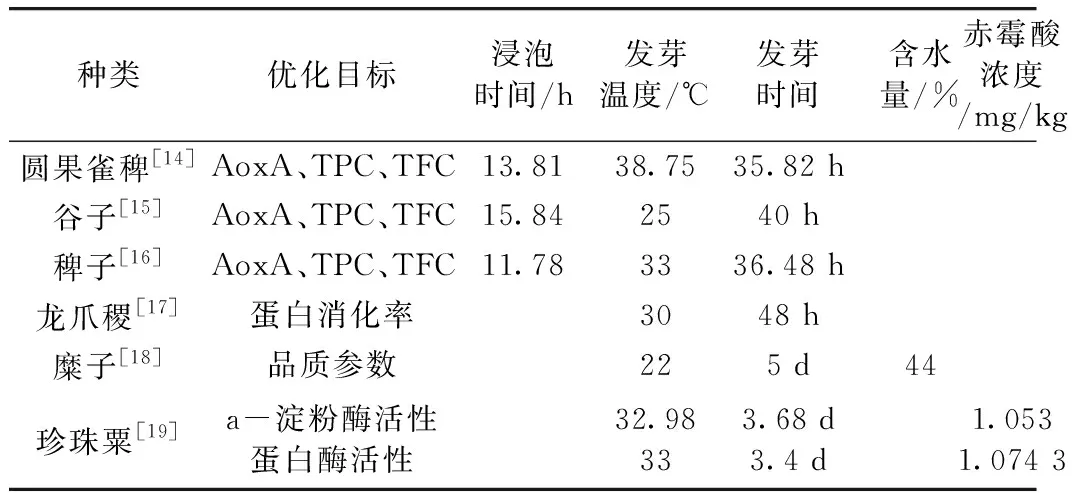
表1 粟米浸泡发芽条件优化

表2 粟米培养基发芽条件优化
注:AoxA:抗氧化活性;TPC:总酚含量;TFC:总黄酮含量;GABA:γ-氨基丁酸;GAD:谷氨酸脱羧酶。
粟芽中的淀粉酶活性不仅与发芽温度及时间有关,也会受到粟米中赤霉酸浓度的影响。赤霉酸是一种由种子发芽产生能够介导酶合成的植物激素,在发芽时撒上不同浓度的赤霉酸溶液可以影响淀粉酶等多种酶的活性。因此为获得更高的淀粉酶活性,常对赤霉酸浓度进行条件优化。另外,针对以培养基为基质的发芽,则以培养基成份及培养环境为参数进行条件优化,为了得到含量较高的γ-氨基丁酸(GABA)时,常对培养基中的谷氨酸、磷酸吡哆醛及钙离子进行优化,谷氨酸脱羧酶(GAD)作用于谷氨酸产生GABA,而磷酸吡哆醛与钙离子则能够改变GAD的活性从而间接影响GABA的生成,结果如表2所示。此外,光照辐射[22],浸泡温度[23],超声处理[24]等条件对粟米发芽也有一定影响。如,γ射线会使发芽率降低,浸泡温度则会影响粟米的吸水率,超声波处理后可溶性蛋白质、γ-氨基丁酸和游离氨基酸含量会有所增加。
粟米品质受到发芽条件及发芽基质的影响,适当地综合考虑这些条件参数就可以显著提高粟米的植物化学性质。因此,为获得更稳定的高品质粟芽,发芽条件的优化值得进一步研究。
2 发芽对粟米营养成分及加工性质的影响
粟米发芽是一种简单的能够提高粟米营养价值的传统食品加工操作,通过体内生物转化过程,粟米中各种酶被激活,从而改变蛋白质、脂肪、碳水化合物等主要营养物质及酚类、黄酮类等植物化合物的含量以及加工性质。
2.1 蛋白质与氨基酸
大部分研究表明,发芽后粟米蛋白质含量有所提高(表3)。如,Swami等[10]发现,龙爪稷发芽24 h后,蛋白质含量从14%增加到17.5%,糜子蛋白质含量在发芽7 d后从12.32%增加到14.30%[25],这可能是由于发芽期间发生的干物质损失所致。一方面,呼吸过程二氧化碳和水从种子中逸出产生糖的损失,水溶性物质的浸出和碳水化合物在发芽过程中的代谢也导致种子的干物质损失。如,糜子在发芽7 d后,干物质损失达到16.5%[25]。另一方面,在发芽过程中,蛋白酶和转氨酶在吸涨后被激活,进行蛋白水解和转氨作用,转化的可溶性肽和氨基酸可以在胚芽内合成蛋白质。然而,一些研究却表明,发芽过程可导致蛋白质总含量降低,可归因于低分子质量含氮化合物在粟米的浸泡和漂洗中有所损失[13]。发芽不仅改变粟米蛋白质的含量,也会影响蛋白质的组成。如,糜子在发芽7 d后白蛋白和球蛋白质量分数从20.3%增加到55.9%,醇溶蛋白从57.8%减少到21.5%,而谷蛋白组分变化较小,可溶性蛋白含量也随着发芽时间的延长而提高[25]。虽然关于发芽粟米蛋白质的研究较多,但主要集中在含量及组成方面,发芽对粟米蛋白质微观结构的影响需要进一步探究。
另外,蛋白质水解后产生大量的氨基酸,从而使粟米氨基酸组成得到改善。已发现在发芽7 d后的糜子中总游离氨基酸从0.086%增加至2.64%,赖氨酸和甲硫氨酸则分别从1.64%,2.0%提高至4.40%和2.12%[25]。如,龙瓜稷发牙4 d后,除谷氨酸,脯氨酸,半胱氨酸,蛋氨酸有明显下降外,其他氨基酸含量都有所提高,其中赖氨酸从3.11%提高至4.17%,在珍珠栗发芽中氨基酸变化也基本相同,其中赖氨酸从2.88%提高至2.96%[26]。Mbithi-Mwikya等[27]也报道,发芽后的龙爪稷氨基酸含量为8.56%,与未发芽(8.58%)的变化不大,但赖氨酸含量从2.58%提高到2.75%。赖氨酸是粟类作物中限制性必需氨基酸,发芽已被用作提高赖氨酸含量,改善粟米氨基酸组成的加工方式。
此外,GABA是一种非蛋白质氨基酸,是中枢神经系统中主要的抑制性神经递质之一,且与食物中一些生理营养功能有关。Kanehira等[28]报道,摄入50 mg含有GABA的饮料可有助于减轻身心疲劳。目前普遍认为,发芽能显著提高粟米中GABA含量。如,Sharma等[29]发现谷子发芽之后GABA含量从0.007 15 g增加到0.038 5 g/100 g,稗子中GABA含量则由0.006 36 g提高至0.035 7 g/100 g。这可能是由于发芽活化的谷氨酸脱羧酶使谷子中的L-谷氨酸发生γ-脱羧作用而导致。Choi[30]报道,在发芽过程中,氨基转移酶在发芽期间变得活跃,导致谷氨酸生成并转化为GABA。粟米储存物质的分解以及蛋白质和其他成分的合成也会导致GABA含量的变化。 发芽谷物能够富集GABA已被广泛证实,但要发挥发芽粟米在其中的优势,还需进一步对条件进行优化。
2.2 脂肪
粟米中存储的脂肪主要存在于胚、糊粉层及果皮中,在发芽过程中脂肪酶可能会降解脂肪以产生游离脂肪酸和甘油,为β-氧化和糖异生以及用于幼苗发育提供底物。如,Hejazi等[31]研究了龙爪稷发芽过程中脂肪及脂肪酸的变化,发现发芽后其脂肪含量有所下降,且在26 ℃下降至最低。其中,油酸(C18∶1N9C)略有下降,在30 ℃发芽48 h后达到最小值43.59%,棕榈酸(C16∶0)和亚油酸(C18∶2N6C)略有增加,亚油酸能够转化为花生四烯酸(ARA)。另外,大部分研究表明,在发芽过程中,脂肪含量会有所下降(表3),这可能是因为在发芽期间脂肪酶的活性和脂肪利用率增加,脂质的水解和脂肪酸在发芽过程中的氧化期间会使脂肪产生损失。也有一些报道显示,发芽期间脂肪含量有所提高,但并未对其原因进行解释,可能为发芽过程中干物质减少所导致。与其他主要成分相比,发芽对粟米脂肪的影响并未得到广泛关注。事实上,粟米糠油富含多种营养成分和生物活性因子,如植物甾醇、维生素E等。对发芽粟米中油脂的深入研究能促进粟米资源的合理应用。
2.3 碳水化合物
粟米中的碳水化合物主要是淀粉,淀粉以直链淀粉和支链淀粉形式储存在胚乳中,发芽过程中淀粉酶活性增强从而水解淀粉以产生六碳糖,导致淀粉含量下降,游离糖含量提高。Sharma[32]发现谷子淀粉酶活性随着发芽时间而提高,并在25 ℃发芽72 h达到最大。龙爪稷在20 ℃下发芽6 d具有最高的α-淀粉酶活性,而在30 ℃下发芽5 d则具有最高的β-淀粉酶活性[9]。Nirmala等[33]发现龙爪稷在发芽四天后,游离糖总量从1.5%提高至16.0%,而淀粉含量则从65%降至43%。在发芽过程中,α-淀粉酶由赤霉酸合成,可从淀粉内部随机水解直链淀粉和支链淀粉的α-1,4键以释放单糖和多糖,葡萄糖和麦芽糖是主要的分解产物。发芽还会改变粟米中淀粉的组成,龙爪稷在30 ℃下发芽48 h后抗性淀粉含量减少了约70%,但可消化淀粉含量基本不变[31]。发芽糜子也有相同趋势,且糜子中的直链淀粉随发芽时间先略有升高,随后迅速减小,在发芽2 d后直链淀粉含量达到最大[25]。但也有少量研究发现,发芽后粟米淀粉含量略有增加,这可能与发芽温度和发芽时间有关。温度过高或过低及发芽时间较短可能使淀粉酶未能充分活化,淀粉分解速率较慢导致含量略微升高。
淀粉是粟米中的主要成分,目前,对发芽粟米中淀粉的研究较为完善。但淀粉并不以单一的组分形式存在于粟米中,进一步探究发芽对粟米淀粉与其他组分(如蛋白质、脂肪、酚类物质等)结合的影响,对深入了解发芽粟米性质变化具有重大意义。
2.4 酚类与黄酮类物质
粟米中酚类与黄酮类物质主要存在于粟米的胚乳及果皮中,且与食品的抗氧化功能密切相关。大多数研究表明,发芽能提高粟米中酚类物质含量,从而提高抗氧化能力。如,Sharma等研究了谷子[29]、稗子[16]、圆果雀稗[14]在发芽过程中总酚及总黄酮含量的变化,发现发芽能提高其含量。这是由于内源酶在发芽过程中被激活并增强了多酚化合物释放,如羟基肉桂酸酯(如阿魏酸和对香豆酸)的多酚化合物通过酯键和醚键等缔合作用与非淀粉多糖结合,而细胞壁降解酶(主要是酯酶)对这些键的作用有助于结合的多酚化合物的释放。然而也有一些研究表明,粟米发芽后酚类物质会有所下降。如,Abdelrahaman等[34]报道发芽6 d的谷子多酚含量下降35%~42%。Subba研究发现粟米在发芽4 d后,原儿茶酸降低了3倍,咖啡酸降低了4倍,而香豆酸,没食子酸和阿魏酸的游离酚酸含量却显著增加,但咖啡酸、香豆酸和阿魏酸的结合酚酸含量下降了2倍[35]。由于粟米中的多酚以游离形式和结合形式存在,由此推测,发芽过程中酚类物质减少可能是由于浸泡水解期间导致多酚物质的损失。除了抗氧化特性外,酚类物质与粟米的感官特性相关,过高的酚类含量可能导致苦味和涩味等不良的口感。
2.5 抗营养因子与矿物质
粟米发芽过程中,抗营养因子如单宁酸和植酸的减少(表4)能够提高矿物质的生物利用度。抗营养物质会结合矿物质,从而影响其生物利用度,随着其浓度降低,其结合能力下降,矿物质变得更容易被利用。Platel等[36]比较了发芽后棕色和白色龙爪稷的矿物质含量,结果表明两个品种的矿物质总量略有下降,但是钙、铁、铜和锰的生物可利用性增加,其中铁的生物利用度增加了4倍。Srinivasan[37]则发现龙爪稷发芽2 d后,随着单宁和植酸含量水平的下降,铁的生物利用度增加了20%。
微量营养素缺乏在发展中国家普遍存在,特别在铁、锌和钙中,发芽已被作为提高植物食品中营养素生物利用度的方法广泛使用。如,Coulibaly等[38]利用发芽3 d后的谷子制备高浓度矿物质面粉。另外,植酸和单宁酸等抗营养物质还是影响淀粉及蛋白质消化率的主要因素。如,Sehgal[39]发现发芽能明显提高珍珠粟的蛋白质(14%~26%)和淀粉(86%~112%)的体外消化率,并且发芽的改善效果显著高于漂烫。抗营养因子能够与淀粉酶、蛋白水解酶相互作用,从而抑制酶的活性,导致消化率降低。但在发芽过程中,抗营养物质、淀粉酶和蛋白酶抑制剂减少,结果导致消化率增加。目前对发芽粟米中抗营养因子的研究主要集中在植酸与单宁酸方面,对其他抗营养因子还有待深入。如,对非淀粉多糖等进一步研究有利于提高发芽粟米的营养价值。
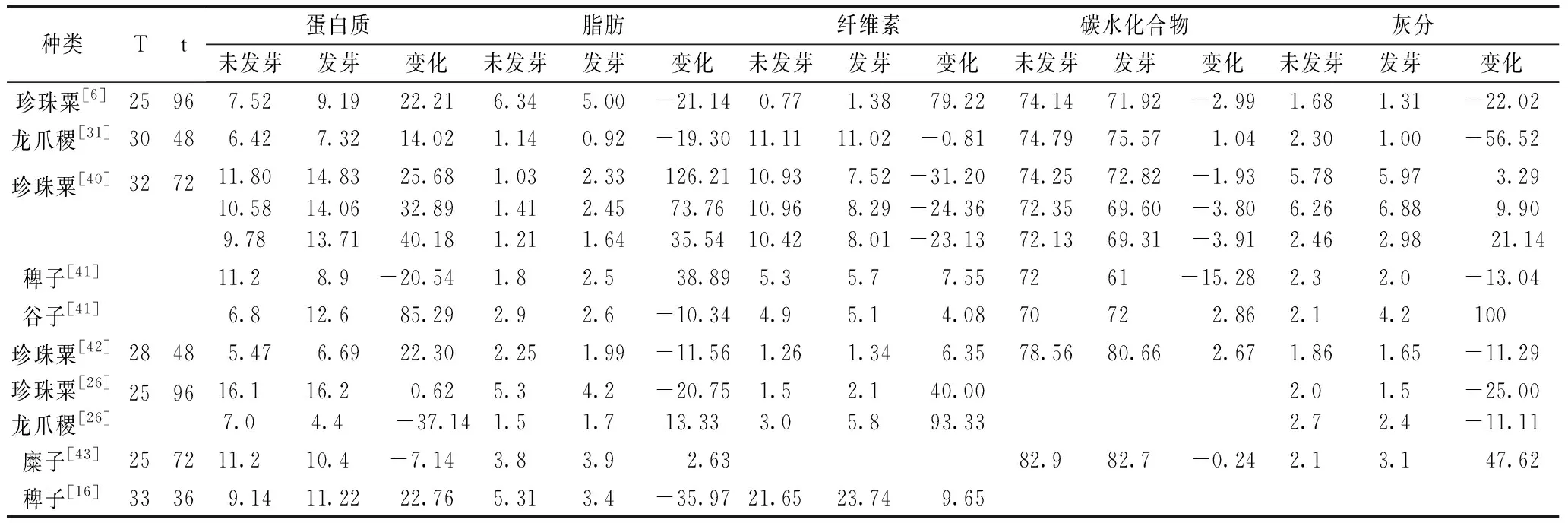
表3 粟米发芽过程中主要成分变化
注:T:发芽温度/℃;t:发芽时间/h;含量变化/%;*含量均以干基表示(g/100 g)。
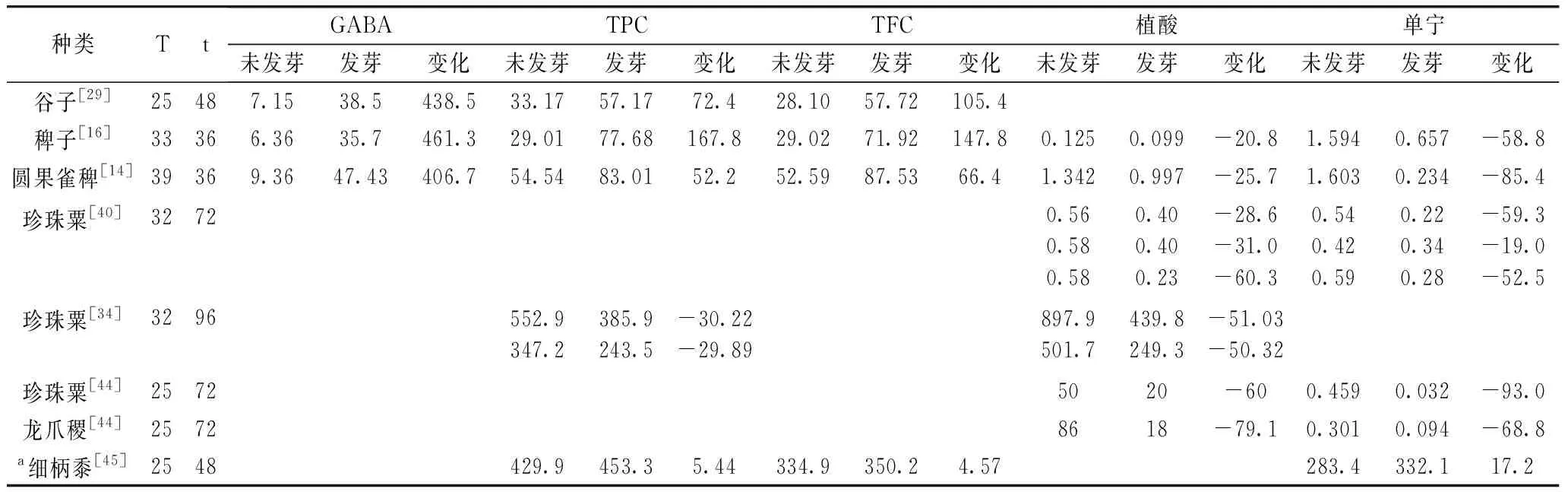
表4 粟米发芽过程中主要功能性成分变化
注:T:发芽温度/℃;t:发芽时间/h;TPC:总酚含量(mg视黄醇/100 g);TFC:总黄酮含量(mg芦丁/100 g);GABA:γ-氨基丁酸(mg/100 g);植酸(mg/100 g);单宁(mol/kg);a:单宁(mg/100 g);*含量均以干基表示。
2.6 发芽对粟米加工性质的影响
物料功能性质与糊化性质是影响粟米加工的主要因素,与最终产品的品质密切相关。Hejazi等[46]研究了龙爪稷发芽之后颜色的变化,发现L值提高,a、b值都有所下降,但随温度及时间变化并没有明显规律。Nazni[41]研究了发芽对稗子和谷子糊化性质(糊化温度,峰值黏度,最终黏度,松懈值,削减值)及物料功能特性(容重、膨胀力、溶解度、固体损失、吸水性和吸油性)的影响。结果表明,除了溶解度和吸油性略有上升之外,其他功能特性指标均有所下降,而糊化特性的所有指标均明显降低,其中糊化温度分别由90.4 ℃和89.6 ℃下降至49.55 ℃和48.60 ℃,其他指标均下降2~40倍。Obadina等[6]研究了发芽对珍珠粟糊化性质及物料功能特性的影响,其结果与稗子及谷子发芽结果一致,且其发现随着发芽时间的延长,糊化温度升高,发芽96 h后其糊化温度达到81 ℃,但仍然小于未发芽糊化温度,而且吸油性和水溶性也随着发芽时间延长而提高。Akinola[47]也证实了发芽后珍珠粟的糊化特性均减小,且发芽后糜子的糊化特性也具有相同的结果[43]。然而Agrawal在研究谷子、稗子和细柄黍的功能性质时发现,发芽会使其吸水性增大[48]Abulub[49]的研究也表明,发芽后粟米的吸水性略有提高,且其起泡性,泡沫稳定性和乳化能力均减小。与之相同,泡沫性质的降低也在珍珠粟发芽中被发现[47]。糊化性质的降低可归因于淀粉分子的连接被发芽期间激活的酶所破坏,从而导致较低的峰值黏度。泡沫性质可能与发芽过程中蛋白质的性质变化有关,在发芽过程中,可溶性蛋白含量增加,结果改善了起泡性。发芽可能导致蛋白质表面变性,并改变分子的表面张力,从而影响泡沫稳定性。吸水性与水溶性也与发芽过程中蛋白质及多糖的含量变化有关,吸油性则与疏水性氨基酸有关。
此外,粟米的加工性质也与发芽过程中的结构变化有关。如,Adebiyi[50]利用X射线衍射,傅立叶红外光谱,扫描电镜观察发芽后珍珠粟的结构,发现其结晶度增加,并且面粉的微观结构从不规则的致密颗粒结构变为规则的松散结构。而Li[51]则发现粟米淀粉的相对结晶度在发芽过程中显著降低,发芽淀粉颗粒表面上出现凹坑和孔洞,而且随着发芽时间增加,颗粒大小分布向小粒径移动,直链淀粉含量降低,但发芽淀粉的分子质量没有显着变化。松散的结构及表面的空洞有利于淀粉酶对淀粉的分解,而较小的颗粒则提高了比表面积,从而提高了分解效率。由于结晶度会影响糊化温度,结晶度高的淀粉不易水解导致较难膨胀糊化,从而使糊化温度提高。目前对发芽粟米加工性质的研究较为粗泛,针对不同产品的加工性质研究尚待进一步细化。
3 发芽粟米在食品中的应用与研发前景
粟米发芽是许多发展中国家的传统加工方式,主要用于生产啤酒,我国传统的谷芽酒就是用谷子发芽后进行酿造。目前我国已有少量粟芽食品的研究,如谷芽大枣汁[52],富集GABA粟芽产品[53]等。而在非洲,粟芽是常用的食品原料,如,Inyang[54]将珍珠粟发芽2 d后制成了fura(一种尼日利亚传统食物),发现发芽的产品在质地和颜色上的评分高于未发芽的产品。Adebiyi等[55]综述了发芽粟米在非洲食品中的应用。就啤酒酿造潜力而言,与高粱相比,珍珠粟芽更具有优势,因为它具有更高的β-淀粉酶活性和更高的游离α-氨基氮含量[56]。另外,发芽龙爪稷可以作为膳食纤维来源,用于制备各种健康食品,且不会改变面团特性或最终产品的质量[57]。此外,发芽提高了能量和营养密度,改善食物的营养成分和消化率,提高了矿物质的生物利用率,使得粟芽成为了婴幼儿食品中的首选的原料之一。发芽也可与其他加工处理(如发酵,挤压,酶解等)组合以提高粟米的营养价值,用于制作各种营养健康的食品,例如面包[58]、饼干[59]、面条[60]及饮料[61]等。另外,粟芽具有天然不含麸质的特性及发芽后的营养强化特点,粟芽在无麸质食品中得到了越来越多的应用。如,无麸质啤酒、饮料以及无麸质面包、饼干[62]等添加粟芽的无麸质食物受到越来越多人的追捧。
发芽作为一种天然的加工方式能够改变粟米的营养性质及加工性质,但受到温度、时间等条件的影响,合理地优化发芽条件能够使粟米发芽朝着有利的方向进行。目前关于粟米发芽后营养成分的变化相互矛盾,尤其是蛋白质、脂肪等主要成分,不同的报道差异较大,可能与不同的发芽条件及粟米种类有关,但粟米发芽能够富集GABA、减少抗营养因子、改善粟米的氨基酸组成已经得到广泛的认可。尽管发芽能够改变粟米的营养性质与功能特性,使其能够适应不同的食品加工,然而,工业规模化应用粟芽还存在一定的困难,主要是能够工业化生产粟芽,且提供高质量粟芽的发芽设备缺乏,而且发芽加工的周期较长也是限制发芽工业化的主要原因之一。因此,开发适应粟米发芽的设备,提供适宜的粟米发芽条件,缩短粟米发芽的周期是未来发芽粟米的重要研究方向。此外,我国虽然有丰富的粟米资源,但目前主要集中在饲料,制粉等初级加工阶段,产品种类较单一,粟米的精深加工非常欠缺,粟米发芽值得进一步研究与开发应用。
