核酸适体结构转换荧光法检测黄曲霉毒素B1
2019-02-15谢岩黎
班 珺 谢岩黎
(河南工业大学粮油食品学院1,郑州 450052)(河南省粮油食品安全检测与控制重点实验室2,郑州 450052)(广西崇左市食品药品监督管理局;崇左市食品药品检验所3,崇左 532200)
黄曲霉毒素是由黄曲霉、特曲霉以及寄生曲霉等产生的一类含有二氢呋喃环结构的次生代谢产物[1-2]。在已分离出的20余种黄曲霉毒素中,黄曲霉毒素B1(Aflatoxin B1,AFB1)致癌性最强,被认为是诱发恶性肿瘤原发性肝细胞癌(hepatocellular carcinoma,HCC)的主要因素之一[3]。为保障食品安全及人类健康,GB 2761—2017[4]对我国食品中AFB1的限量规定为:花生、玉米及其制品≤ 20 μg/kg,稻谷、糙米、大米、植物油脂≤ 10 μg/kg,谷物类、豆类、坚果类、酿造调味品≤ 5 μg/kg。长期以来,科研工作者为AFB1的检测探索了许多方法,常见的传统方法有薄层色谱法[5]、高效液相色谱法[6]、液质联用法[7]、酶联免疫吸附法[8]等,新兴的检测方法有电化学免疫传感器[9]、化学发光免疫法[10]、太赫兹光谱法[11]、核酸适体法[12]等,这些方法的产生极大丰富了AFB1的检测。当前针对AFB1的检测方法众多,每种方法各有其优缺点,仪器检测法灵敏度高、结果较为可靠,但所需仪器昂贵、前处理复杂繁琐,不便于大批量检测;免疫学法检测速度快、操作简单,但所用的抗体制备成本高,保存条件苛刻,批间质量不一,易出现假阳性。因此,开发高效、便捷、灵敏的检测方法成为当务之急。
核酸适体(Nucleic acid aptamer)是采用指数富集配体系统进化技术(SELEX)从体外人工合成的单链核酸文库中筛选出能与靶物质高特异性、高亲和力结合的寡核苷酸片段[13-14]。核酸适体因其靶物质范围广、亲和力高、特异性强等特点,在毒素[15-18]、蛋白[19-22]、激素[23-25]、农药[26-28]、金属离子[29-31]、细胞[32-35]等的检测领域成为重要工具。目前,基于核酸适体的AFB1检测方法,如纳米金法[36]、化学发光法[37]、电化学法[38]等相继报道。
本研究根据核酸适体结构转换信号检测靶分子的原理[39-40],将标记有羧基荧光素(Carboxy Fluorescein,FAM)的AFB1核酸适体与标记猝灭基团(Black Hole Quencher 1,BHQ1)的AFB1核酸适体互补链配对杂交,通过AFB1竞争结合核酸适体的位点使荧光恢复,建立了基于核酸适体结构转换荧光法检测AFB1的方法。
1 材料与方法
1.1 仪器与试剂
Cary Eclopse荧光分光光度计:美国VARIAN公司,实验测定参数:激发波长490 nm,发射波长505~600 nm,激发狭缝10 nm,发射狭缝10 nm。
黄曲霉毒素B1(AFB1)、赭曲霉毒素A(OTA)、雪腐镰刀菌烯醇(DON)标准品:Sigma Aldrich公司;玉米赤霉烯酮(ZEN)标准品:北京普天同创生物科技有限公司;Na2HPO4·12H2O、KH2PO4、NaCl、KCl、MgCl2、CaCl2、HCl、三羟甲基氨基甲烷等:分析纯,天津市科密欧化学试剂有限公司。实验用水均为纯净水:杭州娃哈哈集团有限公司。
实验所用AFB1核酸适体及其互补链均由上海生工生物工程有限公司合成并经HPLC纯化,序列如下:核酸适体1:5′-FAM-GTTGGGCACGTGTTGTCTGTCTCGTGCCCTTCGCTAGGCCCACA-3′;猝灭链1:5′ -ACACGTGCCCAAC-BHQ1-3′; 核酸适体2:5′-GTTGGGCACGTGTTGTCTGTCTCGTGCCCTTCGCTAGG CCCACA-FAM-3′; 猝灭链2:5′-BHQ1-TGTGGGCCTAGCG-3′。
1.2 实验方法
1.2.1 荧光基团标记位置及工作液的选择
以PBS为工作液,比较FAM荧光基团标记核酸适体不同端位对荧光恢复情况的影响,选择荧光恢复率最佳的核酸适体链,以Tris-HCl为工作液测定检测系统的荧光恢复率,比较不同工作液对荧光恢复情况的影响。通过(F-F0)/F0求荧光增长率,其中F0为猝灭状态时的荧光强度,F为加入毒素后的荧光强度。
1.2.2 猝灭链浓度的选择
用工作液将BHQ1标记的猝灭链配制成100、120、140、160、180、200、220、240 nmol/L,分别与200 nmol/L FAM标记核酸适体反应30 min并测定其荧光强度,观察不同浓度互补链对核酸适体荧光基团的猝灭程度。
1.2.3 反应时间的优化
将200 nmol/L核酸适体与猝灭链反应30 min后,向反应体系加入50 ng/mL AFB1并记录其加入反应体系10、20、40、60、80、120 min后的荧光强度,选择最适反应时间。
1.2.4 AFB1的定量检测
用PBS缓冲液将核酸适体2、猝灭链2配制成200 nmol/L,将AFB1母液稀释成0、0.1、1、10、50、100、200、300 ng/mL,在优化好的检测体系中进行标准曲线的建立。400 μL 200 nmol/L核酸适体2与400 μL 200 nmol/L猝灭链2反应30 min后,将800 μL一系列不同浓度的AFB1标准液加入反应体系,充分反应1h后测定其荧光强度,以AFB1浓度为横坐标,荧光强度为纵坐标建立荧光法检测AFB1的标准曲线。
1.2.5 方法特异性考察
将赭曲霉毒素A(OTA)、玉米赤霉烯酮(ZEN)、雪腐镰刀菌烯醇(DON)配制成100 ng/mL。以PBS作为工作液,将200 nmol/L核酸适体2与200 nmol/L猝灭链2反应30 min,分别向反应体系中加入100ng/mL OTA、ZEN、DON,平衡孵育1h后采用荧光分光光度计测定荧光强度。通过(F-F0)/F0对该检测方法的特异性进行分析,其中F0为猝灭状态时的荧光强度,F为加入毒素后的荧光强度。
1.2.6 样品测定
将花生、玉米粉碎,过20目筛,称取5 g样品,以20 mL乙腈-水(85∶15)为提取液高速均质5 min,4 000 r/min高速离心5 min后取4 mL上清液,氮吹仪吹干提取液,加入1mL PBS工作液复溶,采用优化好的检测方法测定样品的初始浓度C0。回收实验中,向样品加入不同量的AFB1,使样品含AFB1分别为5、20、60 μg/kg,采用优化好的检测方法检测加标后的检测浓度C1,以(C1-C0)/加标浓度计算回收率。
2 结果与讨论
2.1 实验原理
检测原理如图1所示,AFB1竞争结合核酸适体的位点,导致猝灭基团标记的核酸互补链脱落,核酸适体构象发生改变,检测体系荧光恢复,根据荧光强度变化对AFB1进行定量检测。检测系统初始状态为标记荧光基团(FAM)的AFB1核酸适体与标记猝灭基团(BHQ1)的互补链通过碱基互补配对结合,由于二者之间距离很近,导致FAM被BHQ1猝灭,检测体系处于弱荧光的猝灭状态。当待测物中含AFB1,AFB1将与核酸适体发生特异性结合,核酸适体结构的改变迫使互补链从核酸适体上脱落,使荧光基团与猝灭基团的距离变远,检测体系中荧光强度得以恢复。当待测物中无AFB1存在时,核酸适体结构不发生变化,荧光强度维持初始的猝灭状态。

图1 AFB1检测原理示意图
2.2 荧光基团标记位置及工作液的选择
实验对FAM荧光基团标记核酸适体端位及反应体系的工作液进行了研究。核酸适体1为5′端标记FAM的寡核苷酸序列,核酸适体2为3′端标记FAM的寡核苷酸序列。图2为等浓度核酸适体1、核酸适体2于不同工作液中的荧光强度比较,图3为核酸适体1、核酸适体2在猝灭状态加入AFB1后的荧光增长率。由图2、图3比较可知,工作液为PBS时,等浓度的核酸适体2荧光强度高于核酸适体1,造成该结果的原因可能为核酸适体5′ 末端的鸟嘌呤对FAM荧光基团产生了一定的猝灭作用,荧光强度较标记在3′端序列的荧光强度弱。当加入猝灭链使FAM猝灭80% 后加入50 ng/mL AFB1,核酸适体2的荧光增长率(60.08%)明显高于核酸适体1的荧光增长率(17.82%),该结果表明FAM标记在3' 端的核酸适体较标记在5' 端的核酸适体更有利于与AFB1结合,这与核酸适体自身形成的空间位阻及AFB1有效识别区域有关。
以核酸适体2进一步研究工作液对荧光强度的影响,由图2可知,等浓度核酸适体下,以PBS为工作液的荧光强度略高于以Tris-HCl为工作液的荧光强度,当加入猝灭链使FAM均猝灭80% 后加入50 ng/mL AFB1,以PBS为工作液的核酸适体荧光增长率(60.08%)高于以Tris-HCl为工作液的荧光增长率(30.58%)(图3)。上述结果表明以PBS为工作液更有利于FAM的荧光发生,也更有利于AFB1与核酸适体的结合。综合考虑,选择核酸适体2并以PBS为工作液进行后续实验探究。

图2 200 nmol/L核酸适体1与核酸适体2于不同工作液中的荧光比较

图3 猝灭状态下加入50 ng/mL AFB1后的荧光增长率
2.3 猝灭链浓度的选择
向200 nmol/L核酸适体2中加入一系列浓度(0、100、120、140、160、180、200、220、240 nmol/L)猝灭链2,充分反应30 min后测定荧光强度如图4所示。猝灭链在100 nmol/L至200 nmol/L时,随着浓度的增加,反应体系中荧光强度迅速降低,当猝灭链浓度为200 nmol/L时,反应体系中的荧光强度已被猝灭80%,继续加大猝灭链浓度,反应体系的荧光强度仍呈减弱状态,但变化趋势向缓。为防止猝灭链浓度过大对AFB1竞争核酸适体的结合位点造成影响,实验最终选择200 nmol/L作为猝灭链最适浓度进行后续研究。

图4 猝灭链浓度的优化
2.4 反应时间的优化
200 nmol/L核酸适体2与200 nmol/L猝灭链2反应30 min后,向反应体系加入50 ng/mL AFB1,图5为AFB1加入反应体系后的10、20、40、60、80、120 min后的荧光强度。由图看出,在AFB1加入后的10至60 min,荧光强度迅速增强,说明随着反应时间的增加,AFB1与核酸适体结合越充分,FAM与BHQ1之间的距离变得越远,荧光强度越强。AFB1加入60 min后,荧光强度较60 min时略微增强但程度趋缓,从检测效率的角度考虑,最终选择60 min为AFB1与核酸适体最适反应时间。

图5 反应时间的优化
2.5 AFB1的定量检测
在优化好的检测体系中对一系列浓度AFB1进行检测,图6为0、0.1、1、10、50、100、200、300 ng/mL AFB1加入反应体系后的荧光光谱图。由图7可知,随着AFB1浓度的增加,FAM与BHQ1距离变远,反应体系由猝灭状态开始恢复荧光,AFB1浓度越大,荧光强度越强。结果表明,AFB1在1~300 ng/mL浓度范围内与反应体系荧光强度具有良好的线性关系,线性方程为Y=0.759 9X+148.928 3(R2=0.996 0),以空白样品均值的3倍标准偏差除以标准曲线斜率计算得到该检测方法的理论检出限为0.8 ng/mL。
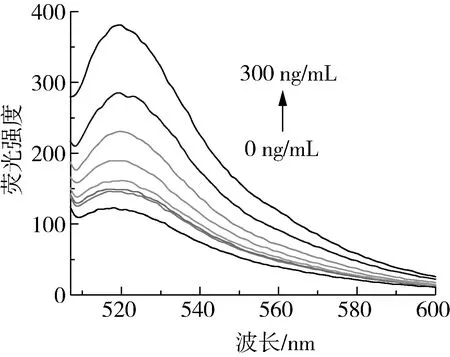
图6 不同浓度AFB1加入反应体系后的荧光光谱图

图7 不同浓度的AFB1检测
2.6 特异性
为考察本方法的特异性,选择粮食中常见的几种毒素:赭曲霉毒素A(OTA)、玉米赤霉烯酮(ZEN)、雪腐镰刀菌烯醇(DON),将它们配制成100 ng/mL并运用本研究建立的AFB1检测方法对其进行检测。实验结果如图8所示,在反应系统中分别加入100ng/mL DON、OTA、ZEN后,荧光增长率分别为0.20%、3.29%、12.68%,与反应猝灭状态时的荧光强度接近,而AFB1在100 ng/mL时的荧光增长率为90.44%,远远高于其他毒素,说明实验所选用的核酸适体对AFB1具有极强的识别选择性,本研究建立的AFB1检测方法特异性较强。

图8 方法选择性考察
2.7 回收率
以花生、玉米为样本向其中添加AFB1标准品,使浓度分别为5、20、60 μg/kg,采用本研究建立的方法进行检测,实验结果如表1所示,花生中的AFB1回收率为95.4%~108.0%之间,玉米中的AFB1回收率为77.7%~84.7%之间,效果良好。

表1 实际样品的测定
3 结论
利用核酸适体能对靶物质特异性结合迫使互补链解离的作用,建立了核酸适体识别AFB1荧光恢复检测法。该法操作简便、灵敏度高、选择性好,可以满足实际样品中AFB1的检测需求。该检测方法的原理同样适用于对其他基于核酸适体检测小分子靶物质的方法设计。
