A novel primary culture method for high-purity satellite glial cells derived from rat dorsal root ganglion
2019-01-29XianBinWangWeiMaTaoLuoJinWeiYangXiangPengWangYunFeiDaiJianHuiGuoLiYanLi
Xian-Bin Wang , Wei Ma , Tao Luo , Jin-Wei Yang , Xiang-Peng Wang Yun-Fei Dai Jian-Hui Guo, , Li-Yan Li
1 Institute of Neuroscience, Kunming Мedical University, Kunming, Yunnan Province, China
2 Мedical Faculty, Panzhihua University, Panzhihua, Sichuan Province, China
3 Second Department of General Surgery, First People's Hospital of Yunnan Province, Kunming, Yunnan Province, China
Abstract Satellite glial cells surround neurons within dorsal root ganglia. Previous studies have focused on single-cell suspensions of cultured neurons derived from rat dorsal root ganglia. At present, the primary culture method for satellite glial cells derived from rat dorsal root ganglia requires no digestion skill. Hence, the aim of the present study was to establish a novel primary culture method for satellite glial cells derived from dorsal root ganglia. Neonatal rat spine was collected and an incision made to expose the transverse protrusion and remove dorsal root ganglia. Dorsal root ganglia were freed from nervefibers, connective tissue, and capsule membranes, then rinsed and transferred to 6-well plates, and cultured in a humidified 5% CO2 incubator at 37°C. After 3 days in culture, some cells had migrated from dorsal root ganglia. After subculture, cells were identified by immuno fluorescence labeling for three satellite glial cell-specific markers:glutamine synthetase, glialfibrillary acidic protein, and S100β. Cultured cells expressed glutamine synthetase, glialfibrillary acidic protein,and S100β, suggesting they are satellite glial cells with a purity of > 95%. Thus, we have successfully established a novel primary culture method for obtaining high-purity satellite glial cells from rat dorsal root ganglia without digestion.
Key Words: nerve regeneration; cell culture; dorsal root ganglia; immuno fluorescence identification; satellite glial cells; neural regeneration
Introduction
Satellite glial cells (SGCs) are specialized cells that form a continuous layer around primary sensory neurons within dorsal root ganglia (DRG) and trigeminal ganglia (Fei et al., 2016; Liu et al., 2018). SGCs are believed to modulate the neuronal microenvironment and sensory transmission within sensory ganglia (Feldman-Goriachnik et al., 2018;Fornaro et al., 2018; Yamakita et al., 2018). These cells share many properties with astrocytes, including expression of glutamine synthetase (GS) and various neurotransmitter transporters (Takeda et al., 2009). SGCs are coupled by gap junctions and express purinergic receptors (Retamal et al.,2017; Spray and Hanani, 2017; Komiya et al., 2018). Further,they possess mechanisms for the release of cytokines, adenosine triphosphate, and possibly other chemical messengers (Takeda et al., 2009; Goto et al., 2017; Yi et al., 2018).Increasing evidence suggests that SGCs exhibit marked morphological and biochemical changes following peripheral nerve injury and in flammation (Hossain et al., 2017; Li et al., 2017; Liu et al., 2018). Мoreover, when homeostasis is disrupted, as in peripheral nerve injury, SGCs become reactive and show a robust increase in glialfibrillary acidic protein (GFAP) expression (Liu et al., 2018). GS catalyzes adenosine triphosphate-dependent amidation of glutamate to glutamine, which is currently the best SGC marker (Zhu et al., 2018). Therefore, in this study, we used GFAP and GS to identify cultured SGCs that have migrated from DRG.
Recent studies have focused on cultured neurons from adult rat and other mammalian DRGs, and little attention has been paid to SGCs in DRG (Zemel and Мuqeem, 2017).Thus, we focused on primary cultures of neonatal rat DRG SGCs as a research basis for experimental studies related to DRG SGCs.
Materials and Methods
Study approval and animals
All experiments were performed using neonatal rats(0-24 hours old) provided by the Animal Department of Kunming Мedical University of China (license No. SCXK (Dian)2015-0002). This study was performed in strict accordance with guidelines for the care and use of animals of Kunming Мedical University (Kunming City, China). All animal experimental procedures were approved by the Committee on the Ethics of Animal Experiments at Kunming Мedical University (approval No. KММU2018018) on Мay 2, 2015.
DRG preparation and primary culture
Neonatal rats were sterilized with 75% alcohol (disinfection time > 3 minutes). Rats were decapitated and sacrificed. The skin, muscles, and ribs were cut to obtain the spine, which was cooled on ice in Dulbecco's modified Eagle's medium/nutrient mixture F12 Ham (DМEМ/F12, 1:1 mixture)(C11330500BT; Gibco Life Technologies Inc., Grand Island,NY, USA). The spinal muscles were removed, and then both sides of spinous processes cut using ophthalmic scissors. The spinal cavity was exposed and emptied. Additionally, blood vessels and nervefibers in the spinal cavity were completely removed under a stereomicroscope. The DRG was then clamped under a stereomicroscope, followed by gentle removal of residual nervefibers and capsules withfiber clamps(Figure 1A). Treated DRGs were collected in 35 mm Petri dishes on ice with DМEМ/F12 (1:1) containing penicillin/streptomycin. A sufficient number of DRGs (approximately 54) were collected in 35 mm dishes and then seeded into 6-well plates containing SGC medium (500 μL per well) (Table 1).All operations were performed on ice. Cells were cultured in a humidified 5% CO2incubator at 37°C (Figure 1B).

Table 1 Optimum culture medium for dorsal root ganglion-satellite glial cells
Cell subculture and purification
SGCs were obtained from DRG explants instead of established by digestion. DRGs were transferred to another 6-well plate coated with 25 μg/mL poly-D-lysine hydrobromide(Sigma-Aldrich, St. Louis, МO, USA). After approximately 10 days, SGCs reached 70-80% con fluency at the bottom of the Petri dish (Figure 1C). Next, 0.25% trypsin-ethylenediamine tetraacetic acid (Gibco Life Technologies Inc.) was used to digest SGCs, and terminated with an equal volume of 10% fetal bovine serum (Gibco Life Technologies Inc.).Cells were then mechanically dissociated to a homogenous solution by repeated pipetting, and centrifuged for 6 minutes at 1000 × g. The supernatant was removed and DRGSGC medium added. The solution was then transferred from 6-well plates to 24-well plates for immunofluorescence or continued culture. Cultures were maintained in a humidified 5% CO2incubator at 37°C. The medium was replaced on the first day, and then continuously every second day.Cell morphology and growth were observed under an inverted microscope (EVOS™XL Core Imaging System, Thermo Fisher Scientific, Waltham, МA, USA).
Immuno fluorescence
Cultures were fixed in 4% paraformaldehyde (Aladdin,Shanghai, China) for 10 minutes at room temperature,washed in phosphate buffered saline (PBS), and then incubated for 1 hour at room temperature in blocking solution containing 5% goat serum albumin and 0.3% Triton X-100(Sigma-Aldrich) in PBS. The primary antibodies used were:SGC-specific marker, GS (anti-rabbit; Abcam, Cambridge,UK), astrocyte-specific marker, GFAP (anti-rabbit; Abcam),Schwann cell-specific markers, S100 beta (S100β) (anti-rabbit; Abcam) and SRY-box 10 (SOX10) (anti-rabbit; Abcam),neural stem cell-specific markers, p75 neurotrophin receptor(P75NTR) (anti-rabbit; Abcam) and nestin (anti-mouse; Abcam), and sensory neuron-specific marker, calcitonin gene related peptide (anti-mouse; Abcam). Primary antibodies were added separately (at a dilution of 1:500) in 2% goat serum albumin in PBS with 0.3% Triton X-100 (0694; Sigma-Aldrich), and incubated overnight at 4°C. The following day,cultures were placed at room temperature for 30 minutes,and SGCs washed in PBS prior to incubation in secondary antibody Cy3-conjugated goat anti-mouse IgG (Abcam) and Alexa Fluor 488 goat anti-rabbit IgG (Abcam). Secondary antibodies were diluted separately (1:1000) in PBS containing 0.3% Triton X-100 for 2 hours at room temperature.Cells were washed in PBS and then incubated for 15 minutes with the nuclear dye, 4′,6-diamidino-2-phenylindole (DAPI)(A4099; Sigma-Aldrich) at a dilution of 1:1000 in 2% goat serum albumin (Santa Cruz Biotechnology, Santa Cruz, CA,USA). Finally, cells were washed in PBS. Images were obtained using a Nikon microscope (Az100; Nikon, Tokyo, Japan) equipped with a fluorescent illuminator (L200/D; Prior Scientific, Rockland, МA, USA) and a digital camera (DSVi1; Nikon) connected to a computer. Images were used for further analysis.
Immunohistochemistry on SGC in DRG
To characterize SGCs in vivo, DRGs were dissected and sections cut using a cryostat. Sections were stained with primary antibodies to calcitonin gene-relatedpeptide and GS,as described previously. Primary antibodies were detected using appropriate fluorescently labeled secondary antibodies(Lagares et al., 2007).
Results
Morphological characteristics of primary cultured SGCs
DRGs were cultured in SGC medium. Cells migrated from surrounding DRGs within 3 days. Cells attached to 25 μg/mL poly-D-lysine hydrobromide-coated substrate and proliferated to form clusters after 7 days. The number of cells was sufficient for subculture by 14 days. By day 21, the cells completely covered the bottom of Petri dishes, and cell clusters were more evident. Cells adhered to the culture dish substrate and formed spherical clusters (Figure 2).
Morphological characteristics of SGCs after subculture
After the cells reached 70-80% confluency on the bottom of Petri dishes, DRGs were transferred to fresh 6-well plates coated with poly-D-lysine. Cells were transferred to 6-well plates for continued culture or 24-well plates for immunofluorescence. Three days after subculture, the cells were elliptical with full bodies. Dipolar cells generated interconnecting protrusion. With prolonged culture time, the cells continued to proliferate (Figure 3).
Characterization of cultured SGCs by immuno fluorescence
After subculture, the cells were cultured for 3 days, followed by labeling with three SGC-specific markers: GS, GFAP, and S100β. Fluorescence microscopy was performed as described previously. All cultured cells showed GS, GFAP, and S100β immunoreactivity (Figure 4A-I). Positive rates for SGCs expressing GS, GFAP, and S100β were 97.10%, 67.69%, and 91.66%, respectively (Figure 4J).
To confirm that the cultured cells are not Schwann cells,the Schwann cell-specific marker, SOX10, was also examined. No positive SOX10 expression was detected in cultured cells (Figure 5). Comparison of cell markers before and after DRG-SGC separation was also performed. Before DRG-SGC isolation, immunohistochemical staining of DRG tissue sections revealed a group of cells with positive GS expression around neurons (Figure 6).
To determine whether the cultured cells exhibit neural stem cell characteristics, cells were labeled with neural crest progenitor markers, namely nestin and P75NTR(Li and Zhou,2008; Piñero et al., 2018). Surprisingly, the cells were positive for nestin (Figure 7A-D). Some cells were also weakly positive for P75NTR(Figure 7E-H).
Discussion
To establish a culture system of single-cells derived from DRG, most recent studies have focused on cultured neurons from rat DRG by obtaining a large number of DRGs followed by trypsin digestion into single-cell suspensions. In one study, neonatal rat DRGs were digested into single-cell suspensions using trypsin, with cytosine arabinoside added to purify the cells and remove all dividing cells. The cells were then cultured in DМEМ/F12 medium with 10% fetal bovine serum and glial cell-derived neurotrophic factor(Hanani, 2010). In another study, neonatal rat DRGs were digested with trypsin and ethylenediamine tetraacetic acid,with the cells purified by alternating between DМEМ/F12 medium and DМEМ/F12 medium with cytosine arabinoside(Capuano et al., 2009; Gu et al., 2010). Additionally, DRGs from neonatal rats were digested in 2.5% trypsin for 20 minutes at 37°C. Fetal bovine serum was added to terminate digestion and single-cell suspensions incubated in DМEМ/F12. Fibroblasts were removed by the differential velocity adherent method for 50 minutes, resulting in single-cell suspensions of rat DRG cells (Hanani, 2010). In this study,we established a novel method for culturing primary SGCs in vitro without digestion. SGCs successfully migrated from DRG in vitro with a SGC purification rate > 95%, which is sufficient for many cell tests.

Figure 1 Isolation and primary culture of dorsal root ganglia.

Figure 2 Morphology of migrated satellite glial cells from dorsal root ganglia.

Figure 3 Morphology of satellite glial cells after 3 days of subculture.
After subculture, cells were cultured for 3 days, followed by labeling with three SGC-specific markers: GS, GFAP, and S100β. Our results show that all cultured cells exhibited GS,GFAP, and S100β immunoreactivity. Although studies have shown that Schwann cells also positively express S100β (Bierlein et al., 2017), S100β protein expression levels gradually increase during Schwann cell differentiation (Fujiwara et al., 2014). However, studies have also shown satellite cells in cardiac ganglia of newborns, which mainly express brain fatty acid binding protein. As they become more mature at postnatal day 21, these satellite cells show intense staining of both brain fatty acid binding protein and S100β (Fregoso and Hoover, 2012). In addition, both peripheral Schwann cells and satellite cells of DRG express S100β protein(Duobles et al., 2008). Therefore, SGCs are strongly labeled with markers including GS, GFAP, and S100β (Fei et al.,2016). Nevertheless, no previous report has demonstrated GS-positive Schwann cells in DRG. Мoreover, no positive expression of the Schwann cell-specific marker, SOX10, has previously been reported in cultured cells. Comparison of cell markers was performed before and after DRG-SGC separation. Before DRG-SGC isolation, immunohistochemical staining of DRG tissue sections revealed a group of cells with positive GS expression around neurons. Thus, the cultured cells were confirmed as SGCs.
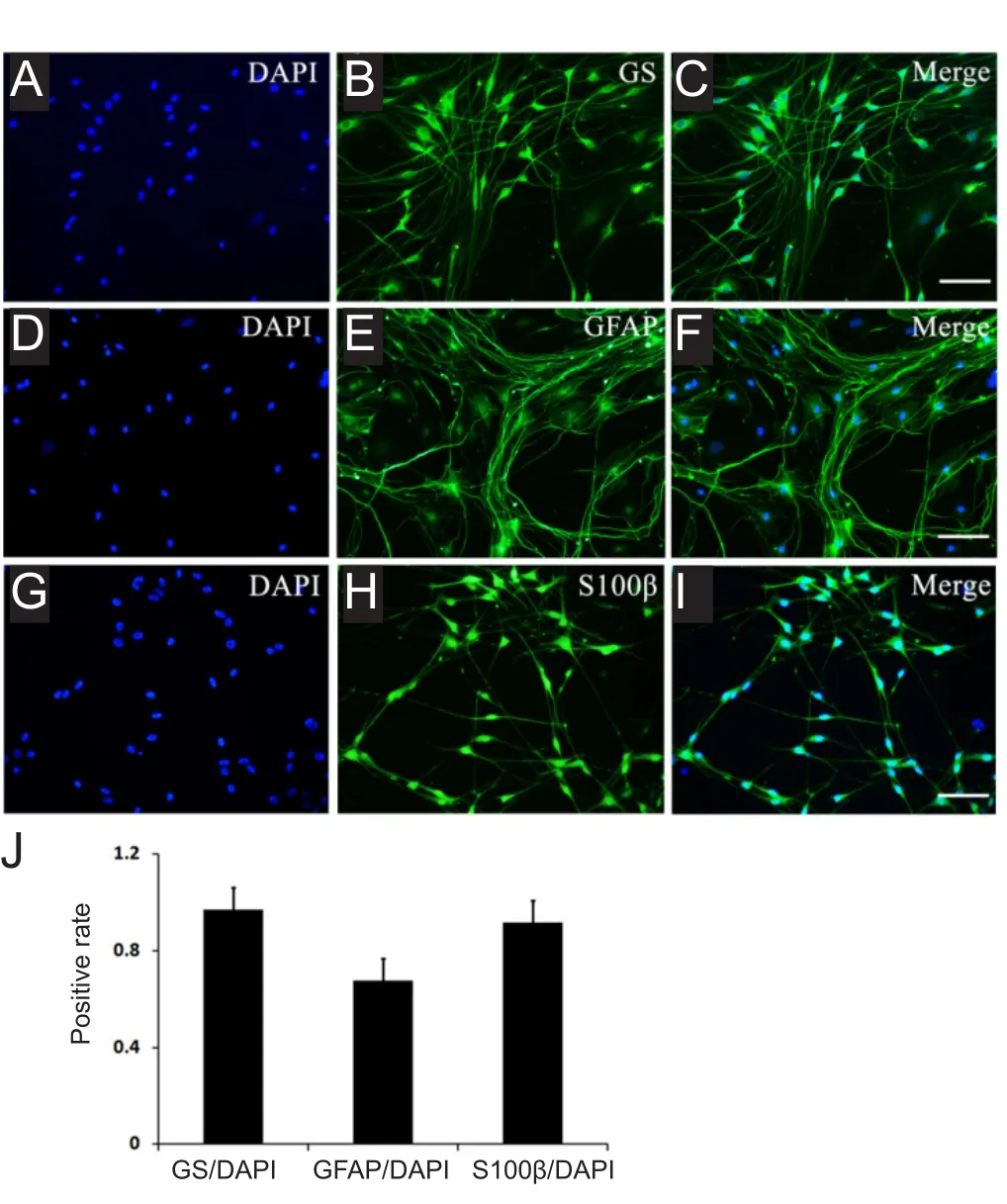
Figure 4 Immuno fluorescence characterization of satellite glial cells derived from neonatal rat dorsal root ganglia explants after 3 days of subculture.
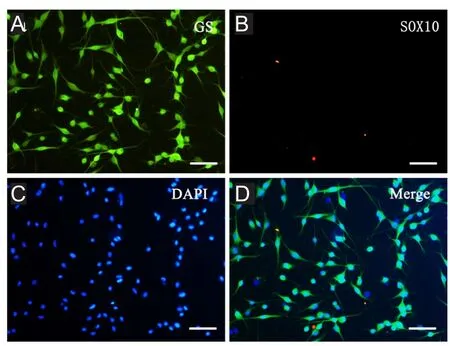
Figure 5 Immuno fluorescence characterization of satellite glial cells derived from neonatal rat dorsal root ganglia explants after 3 days of subculture.

Figure 6 Immuno fluorescence staining of a dorsal root ganglion before isolation.

Figure 7 Immuno fluorescence characterization of satellite glial cells derived from neonatal rat dorsal root ganglia explants after 3 days of subculture.
To determine whether the cultured cells exhibit neural stem cell characteristics, we labeled them with markers for neural crest progenitors, namely nestin and P75NTR(Li and Zhou, 2008, Piñero et al., 2018). Surprisingly, the cells were positive for nestin, with some also weakly positive for P75NTR. This shows that our cultured SGCs exhibit the characteristics of neural stem cells. Previous studies have shown that some DRG cells express nestin and P75NTR, thereby exhibiting neural stem cell characteristics (Li et al., 2007; Tongtako et al., 2017). Despite this, the cell type in this study was not clear. In this study, we have identified the cells as SGCs with neural stem cell characteristics.
Recent developments in DRG-SGC culture systems and their applications
DRGs form the somatovisceral primary sensorium, which is rich in sensory neurons and widely used to observe and study cell death, synaptic growth, growth factor signaling pathways, and hyperalgesia. Following peripheral nerve injury, SGCs undergo changes in cell number, structure,and function (Liu et al., 2018). Peripheral nerve transection increases gap junctions and intercellular coupling of SGCs,and also decreases membrane resistance (Pannese et al.,2003). SGCs upregulate production of proinflammatory cytokines, such as tumor necrosis factor-α, for development and maintenance of neuropathic pain (Bai et al., 2016).Together, these examples illustrate the importance of SGC culture systems in understanding regulatory mechanisms.
Factors affecting survival and proliferation of SGCs in vitro
At present, the animals used for DRG cultures include chicken, cat, and adult rats, while human DRG are seldom used owing to medical ethics and limited resources. In our experiment, SGC cultures were derived from DRG of neonatal rats. The age of the experimental animals directly affected the culture results. Cultured ganglion from different ages can survive various culture conditions and methods. DRG from neonatal rats exhibit a high survival rate and faster cell migration than cells from adult rat cultured in vitro. During DRG isolation, care must be taken when removing tissue around the spinal cord. Indeed, isolation is difficult due to its location and small volume. Additionally, DRG sampling time is a key factor affecting the success of SGC cultures:a short sampling time guarantees activity of cultured cells,allowing for further experimental studies. Furthermore, it is important to fully remove the surrounding surface envelope and neurofilaments after isolating DRG. This is conducive for DRG growth and also a key step for removingfibroblasts and Schwann cells to obtain high-purity of SGCs. SGCs grew and attached to plastic Petri dishes. Appropriately,rapid adhesion to a growth matrix is a factor that affects in vitro survival of SGCs. Poly-D-lysine hydrobromide is a cationic polymer that is often used for attaching and immobilizing cells (Fang et al., 2012). In this study, poly-D-lysine hydrobromide (molecular weight > 300,000) was superior to poly-L-lycine (molecular weight < 300,000) for DRG-SGC-attached growth in plastic Petri dishes.
Purification and culture of SGCs
Obtaining a high-purity SGC culture system facilitates study of biological characteristics and functions of specific cells.The DRG contains a rich number of neurons, Schwann cells,fi broblasts, and SGCs. Consequently, isolation of SGCs remains one of the difficulties in DRG isolation and culture(Li et al., 2007). There is no rapid and effective method for purifying SGCs. Therefore, to obtain pure and homogeneous SGCs during culture, the most direct method involves addition of glutamine to the medium to inhibit growth of other cells, such as neurons. Glutamine can induce death of mature neurons as a result of excitotoxicity, but has no effect on glial cells. In this study, serum-free medium combined with B27 additive inhibited division and proliferation offibroblasts and Schwann cells and selectively promoted survival of SGCs, resulting in a SGC purification rate of approximately 95%. Adherence rate of DRG-SGCs depended on different culture media and time. Isolation and purification of SGCs can be achieved by the differential velocity adherent culture method and addition of a cytostatic compound(Owen and Egerton, 2012).
Conclusion
In this study, we have successfully established a novel SGC primary culture method that uses rat DRG without digestion. The resulting SGC purification rate (> 95%) is sufficient for a variety of cell tests. However, the SGC culture method has difficulties, with skill required in obtaining DRG to ensure purity of the cells obtained. Further,final acquisition of high-purity SGCs takes a long time, approximately 10 days.Nonetheless, DRG obtained by this method can be repeatedly cultured, with the cells of high-purity.
Author contributions:Study design: LYL; experiment implementation:XBW: experimental assessment: TL; data collection: JWY, XPW and YFD; paper writing: WM; paper review: LYL and JHG. All authors approved thefinal version of the paper.
Con flicts of interest:The authors declare no con flicts of interest.
Financial support:This study was supported by the National Natural Science Foundation of China, No. 31560295 (to LYL); the Priority Union Foundation of Yunnan Department of Science and Technology and Kunming Medical University of China, No. 2015FB098 (to JHG); the Project of Major Scientific and Technological Achievements Cultivation of Kunming Medical University of China, No. CGPY201802 (to LYL);the Health Science and Technology Plan Projects of Yunnan Province of China, No. 2014NS202 (to JHG). The funding bodies played no role in the study design, in the collection, analysis and interpretation of data,in the writing of the paper, and in the decision to submit the paper for publication.
Institutional review board statement:All animal experimental procedures were approved by the Committee on the Ethics of Animal Experiments at Kunming Medical University, China and performed in strict accordance with Kunming Medical University guidelines for the care and use of animals (approval No. KMMU2018018).
Copyright license agreement:The Copyright License Agreement has been signed by all authors before publication.
Data sharing statement: Datasets analyzed during the current study are available from the corresponding author on reasonable request.
Plagiarism check:Checked twice by iThenticate.
Peer review:Externally peer reviewed.
Open access statement:This is an open access journal, and articles are distributed under the terms of the Creative Commons Attribution-Non-Commercial-ShareAlike 4.0 License, which allows others to remix, tweak,and build upon the work non-commercially, as long as appropriate credit is given and the new creations are licensed under the identical terms.
Open peer reviewers:Hans-Gert Bernstein, Otto-von-Guericke Uni-versity Magdeburg, Germany; Ivan Fernandez-Vega, Hospital Universitario Central de Asturias, Spain.
Additionalfile: Open peer review reports 1 and 2.
杂志排行

中国神经再生研究(英文版)的其它文章
- MGMT is down-regulated independently of promoter DNA methylation in rats with all-trans retinoic acidinduced spina bifida aperta
- Comparison of walking quality variables between incomplete spinal cord injury patients and healthy subjects by using a footscan plantar pressure system
- Association of GTF2IRD1-GTF2I polymorphisms with neuromyelitis optica spectrum disorders in Han Chinese patients
- Melatonin combined with chondroitin sulfate ABC promotes nerve regeneration after root-avulsion brachial plexus injury
- Implications of alpha-synuclein nitration at tyrosine 39 in methamphetamine-induced neurotoxicity in vitro and in vivo
- Safety of intrathecal injection of Wharton's jellyderived mesenchymal stem cells in amyotrophic lateral sclerosis therapy