鸭甲型肝炎病毒的遗传和抗原稳定性分析
2019-01-14马秀丽李玉峰于可响刘存霞亓丽红宋敏训
马秀丽,黄 兵,李玉峰,于可响,刘存霞,胡 峰,亓丽红,宋敏训,艾 武
(山东省农业科学院家禽研究所 山东省禽病诊断与免疫重点实验室,济南 250023)
鸭甲型肝炎病毒(Duck hepatitis A virus,DHAV)归属于小RNA病毒科禽肝炎病毒属,DHAV主要危害3周龄以下的雏鸭,死亡率高达80%以上,给养鸭业造成巨大危害[1]。DHAV包括三个血清型,即DHAV-1,DHAV-2和DHAV-3[2]。DHAV-1的危害普遍存在于全国各地,近年来,DHAV-3的流行有增加的趋势,个别鸭场还会出现DHAV-1和DHAV-3混合感染的现象,而DHAV-2仅在台湾发生过[3]。三者均具有典型小RNA病毒的基因组结构,但三者之间存在显著的序列差异,遗传进化关系表明三者属于三个不同的基因型,即基因A型、基因B型和基因C型,血清学试验证明三者之间无交叉反应[4,5]。随着DHAV-1和DHAV-3的危害日趋严重[6-15],该病的遗传变异和抗原稳定性备受大家的关注。为进一步分析当前DHAV的流行特点和变异现状,本研究对本实验室保存的25株DHAV-1(2000年~2017年)和32株DHAV-3(2008年~2017年)以及Genbank中下载的DHAV代表株的VP1基因进行了生物信息学分析,结合代表毒株的生物学特性,进一步揭示了DHAV-1和DHAV-3的抗原特性和变异情况,对我国今后鸭甲型肝炎病毒的综合防控具有重要的指导意义。
1 材料与方法
1.1 DHAV毒株和阳性血清本实验室分离保存的25株DHAV-1(表1)和32株DHAV-3(表2)均由本实验室分离保存;1-CL/03、1-HA5/13、3-FX/08和1-HC5/13单因子阳性血清由本实验室制备并保存。
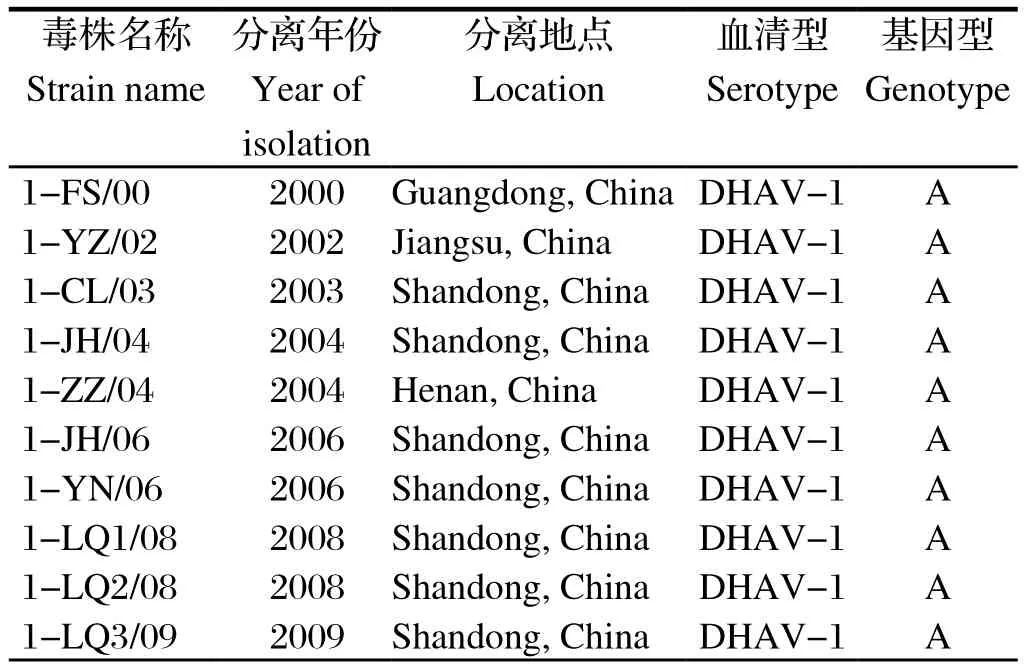
表1 DHAV-1毒株背景Table1 DHAV-1 strains used in this study
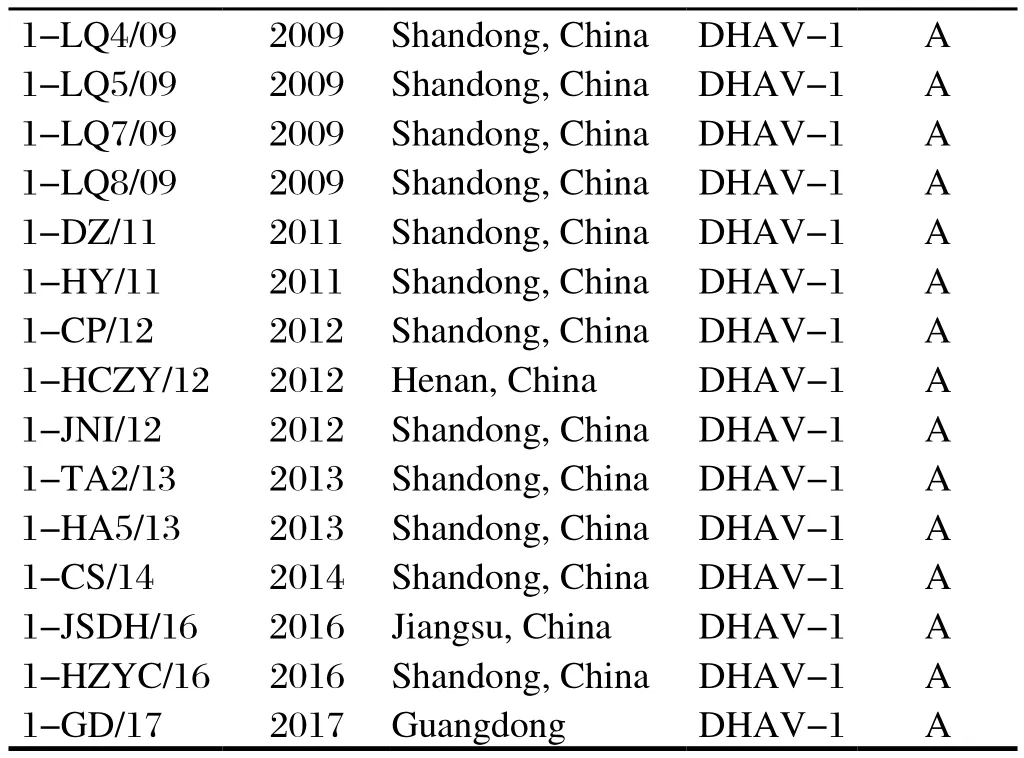
(续上表)

表2 收集的DHAV-3毒株背景Table2 Summary of DHAV-3 strains used in this study
1.2 DHAV VP1基因的扩增参考Genbank中已发表的DHAV-1和DHAV-3 VP1基因序列,分别设计1对扩增DHAV-1、DHAV-3VP1基因异性引物。DHAV-1-F:5'- CTCGAGGGTGATTCTAACC AGTT-3',DHAV-1-R:5'-GCGGCCGCTTCA ATTTCCAGATT- 3'(下划线碱基为酶切位点);DHAV-3-F:5'- CTCGAGGGTGATTCCAATC AGCT- 3',DHAV-3-R:5'- GCGGCCGCTTC AATYTCCARAT- 3'(下划线碱基为酶切位点)。采用Trizol方法提取病毒RNA后,按常规RT-PCR方法扩增DHAV毒株的VP1基因,并将回收的PCR产物克隆至pMD18-T载体中,双酶切法筛选出阳性重组子,送北京博尚生物有限公司进行VP1基因序列测定。
1.3 DHAV VP1蛋白的遗传进化分析采用MEGA6.0软件对测定的DHAV毒株与Genbank中下载的VP1基因(表3)的核苷酸和氨基酸序列进行比较,用Boot-strap法(Replication值为1000)计算遗传距离并绘制N-J进化树,分析当前国内DHAV的流行情况及不同毒株氨基酸的变异程度。
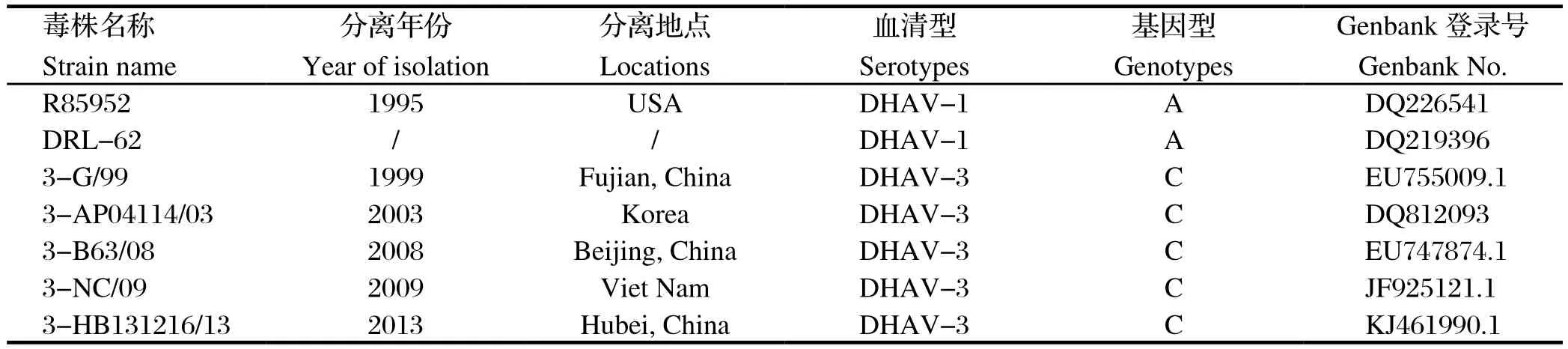
表3 Genbank中下载的DHAV-1和DHAV-3序列Table 3 Reference sequences of DHAV-1 and DHAV-3 from Genbank
1.4 血清交叉中和试验采用固定病毒稀释血清方法进行交叉中和试验。将自制的1-CL/03、1-HA5/13或3-FX/08、3-HC5/13单因子血清分别与等体积的病毒含量为200ELD50/0.1mL的DHAV代表毒株(2016年和2017年)混合,充分作用后分别接种非免鸭胚,37℃温箱培养,记录结果,用Reed和Muench两氏法计算血清中和指数。
2 结果
2.1 DHAV VP1基因的序列分析对实验室DHAV-1、DHAV-3毒株的VP1基因序列分析发现,DHAV-1间的VP1基因的核苷酸序列相似性为91.7%~99.9%;DHAV-3间的VP1基因的核苷酸序列相似性为90.7%~99.6%;DHAV-1和DHAV-3间的VP1基因核苷酸序列相似性为70.0%~73.5%;因此VP1蛋白序列同源性在DHAV分型上起主要作用。DHAV-1和DHAV-3的VP1存在一些氨基酸变异位点,主要集中在143~147位,171~201位,204~212和217~224位。氨基酸变异位点的存在导致了DHAV-1和DHAV-3抗原性的差异,从而使得DHAV-1与DHAV-3的交叉反应性很低或无交叉中和反应[3,5]。选取DHAV-1代表株1-CL/03和1-HA5/13,与1-JSDH/16、1-HZYC/16和1-GD/17分离毒株之间比较,结果发现存在5处氨基酸位点的改变(181位、184位、193位、219位和238位)。选取DHAV-3代表株3-FX/08和3-HC5/13,与2016 ~2017年DHAV-3分离毒株进行比较,发现存在9处氨基酸位点的改变(21位、48位、159位、188位、195位,196位、205位、239位和240位),结合血清中和试验结果,表明这些氨基酸位点的改变没有影响DHAV-1或DHAV-3的中和特性。
2.2 遗传进化分析根据绘制的系统发育树可以看出,DHAV-1进化为两个大分支(图1),一个分支为2013年以前的分离毒株,另一分支为近几年的分离毒,其中2013年的分离毒1-HA5/13也分布在这个分支上,表明2016~2017年分离毒与1-HA5/13遗传距离较近。DHAV-3也进化为2个大分支(图2),一个分支为经典的毒株3-AP04114/03、3-B63/08和3-NC/09,另一个分支为实验室2008~2017年分离毒株和福建分离毒株3-G/99,其中实验室分离毒又进化为一单独的亚分支。
2.3 DHAV血清交叉中和试验由图3可见,3株DHAV-1分离株(1-JSDH/16、1-HZYC/16、1-GD/17)针对1-CL/03和1-HA5/13血清的中和指数均在100以上,均可判为阳性。3株分离毒与1-CL/03和1-HA5/13血清的中和指数差异较大,可能与这2份血清自身效价差异较大有关。由图4可见,10株DHAV-3分离株针对3-FX/08和3-HC5/13血清的中和指数均在100以上,均可判为阳性,表明各毒株之间有较好的交叉反应。
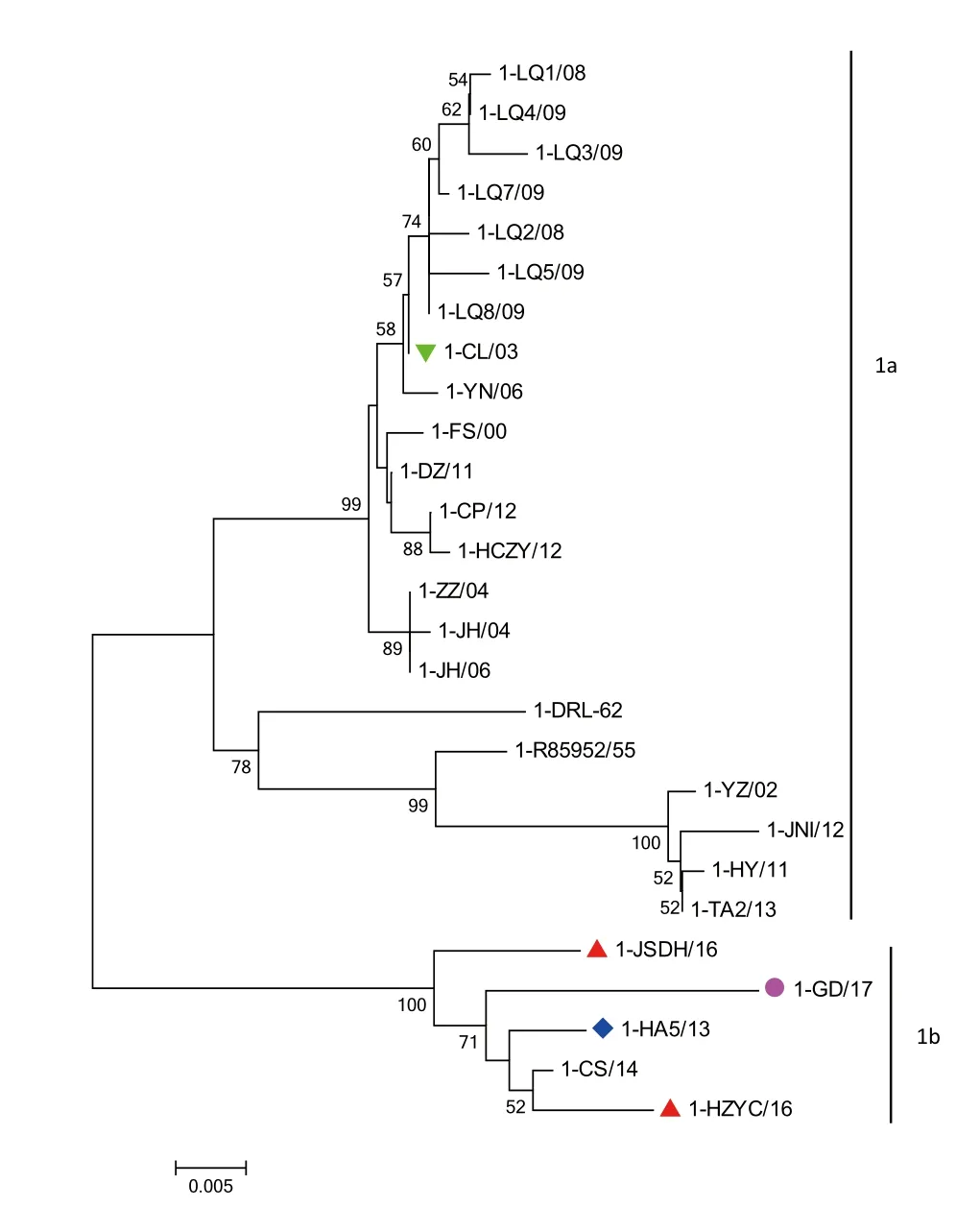
图1 DHAV-1遗传进化关系分析Fig.1 Genetic and phylogenetic analysis of DHAV-1

图2 DHAV-3遗传进化关系分析Fig.2 Genetic and phylogenetic analysis of DHAV-3
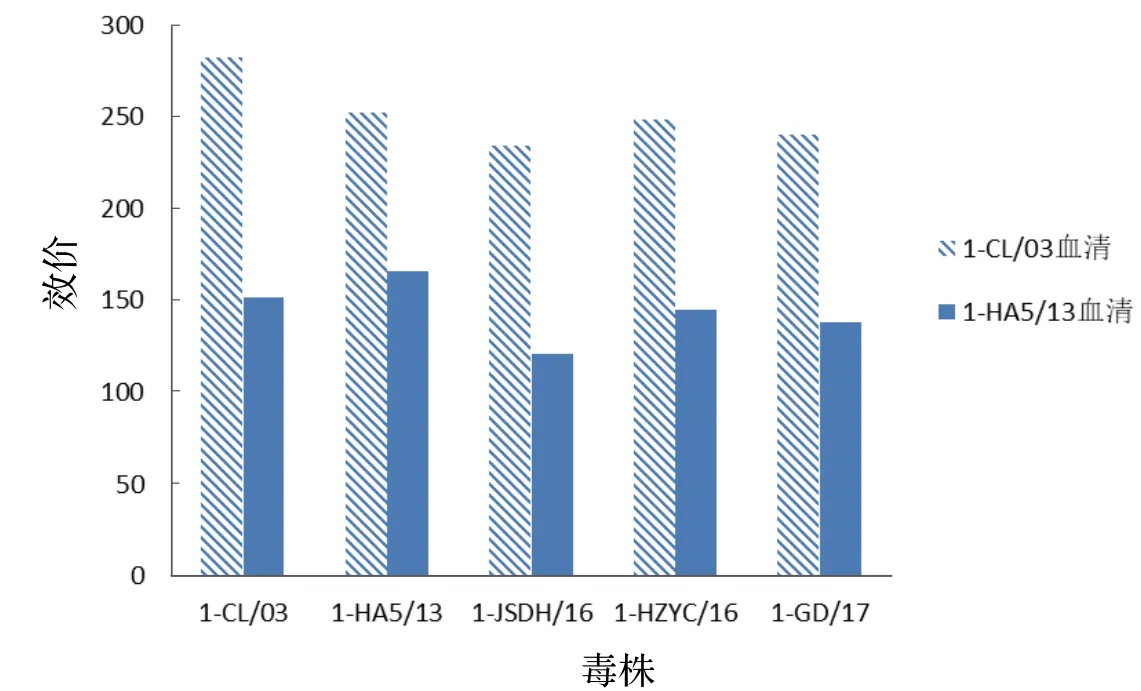
图3 1-CL/03和1-HA5/13针对DHAV-1不同毒株的中和指数Fig. 3 Neutralization index of different DHAV-1 strains against serum 1-CL/03 and 1-HA5/13

图4 3-FX/08和3-HC5/13针对DHAV-3不同毒株的中和指数Fig. 4 Neutralization index of different DHAV-3 strains against serum 3-FX/08 and 3-HC5/13
3 讨论
3.1 DHAV遗传进化分析本文根据DHAV-1和DHAV-3 VP1基因的遗传进化分析发现,在过去16年内,DHAV-1进化为两个主要分支,分支1a包括2000~2013年流行的20个毒株,另一个分支1b包括2013~2017年流行的5个毒株。我们又进一步分析这两个分支上DHAV-1的地理分布差异,结果发现,来源于同一地区的毒株在两个分支中均存在,这可能与当前家禽频繁的贸易往来有较大关系。过去8年内DHAV-3进化为两个遗传分支,其中分支3b只有2003~2009年分离的3个毒株,1个毒株来自韩国(3-AP04114/03),1个毒株来自越南(3-NC/09),1个毒株来自中国北京(3-B63/08)。另一个分支3a为本研究中分离的DHAV-3毒株与福建毒株3-G/99,DHAV-3分离毒株占据单独的一个亚分支,有趣的是这些毒株又进一步分化成更小的亚分支,其中2012-2017年的大多数毒株集中在一个亚分支上。另一个亚分支上只有福建毒株3-G/99,由此可见我国早在1999年就有DHAV-3的存在。DHAV-3毒株之间无明显的地理分布差异。
3.2 DHAV-1和DHAV-3的血清学特征我们选取DHAV-1和DHAV-3两个分支上的代表毒株制备的阳性血清1-CL/03、1-HA5/13、3-FX/08和3-HC5/13,分别与近两年分离的DHAV-1和DHAV-3毒株进行血清交叉中和试验,其主要目的是分析DHAV遗传多样性对抗原性的影响。结果表明,过去16年的DHAV-1和8年的DHAV-3的抗原性相对稳定,相同血清型之间有很好的交叉保护,DHAV疫苗仍能中和当前流行的相同血清型的DHAV毒株,说明现有DHAV疫苗完全能够保护相同血清型的DHAV的感染。
总之,在过去16年中,DHAV的遗传性和抗原性相对稳定。尽管DHAV-3的进化速度远快于DHAV-1[16],目前来看DHAV-3的高进化速率并未造成该血清型病毒抗原性的改变。但上述研究结果提示我们应做好DHAV的流行病学监测,防止潜在的遗传变异导致病毒逃逸而引发疫病的暴发和流行。
