Investigation of cryptic diversity and occurrence of echinostome metacercariae infection in Anentome helena (von dem Busch, 1847)
2018-11-14NithinanChomchoeiChalobolWongsawadNattawadeeNantarat
Nithinan Chomchoei, Chalobol Wongsawad, Nattawadee Nantarat✉
1Department of Biology, Faculty of Science, Chiang Mai University, Chiang Mai 50200, Thailand
2Graduate School, Chiang Mai University, Chiang Mai 50200, Thailand
3Applied Parasitology Research Laboratory, Department of Biology, Faculty of Science, Chiang Mai University, Chiang Mai 50200, Thailand
4Environmental Science Research Center, Faculty of Science, Chiang Mai University, 50200, Thailand
5Science and Technology Research Institute, Chiang Mai University, Chiang Mai 50200, Thailand
Keywords:Phylogeny Freshwater snail Echinostomes Biodiversity Anentome
ABSTRACT Objective: To investigate the parasitic infection of Anentome helena (A. helena) and determine the validity of species boundaries for A. helena by combining molecular phylogeny and morphological approaches. Methods: A total of 325 individuals of A. helena were collected throughout northern Thailand. Shells were measured and compared by t-test. Radulae were investigated by using light and scanning electron microscope. Two partial mitochondrial DNA sequences of COI and 16S rRNA from 36 specimens of A. helena and related species were used to test the validity of the morphospecies. Phylogenetic trees were constructed using neighbour joining, maximum likelihood and Bayesian inference. Infection of A. helena with trematode larva was examined and observed. Results: Morphological examination of A. helena revealed 2 distinct morphospecies. Genetic divergences supported the separation of the two morphotypes into two distinct groups. Both individual and combined analyses of the two nucleotide fragments revealed two phylogroups that corresponded with shell and radula characteristics. In addition, A. helena was found infected with 37-collar spined echinostome metacercariae. The prevalence and intensity of metacercariae was highest in the San Kamphaeng district, Chiang Mai province, with 7.5% and 1.670±0.577, respectively. Conclusions: These findings suggest that comprehensive taxonomic revision of this unrecognised species complex is needed.This study represents the preliminary step to reveal new data on the recent distribution of trematode infection in A. helena. This information may be useful for developing conservation management of the snail and the practice of targeted regimes to reduce anthelmintic resistance in the future.
1. Introduction
The genus Anentome contains carnivorous snails in the Family Nassariidae, one of the largest and most biologically successful families worldwide. This genus consists of 6 species: Anentome helena(A. helena), Anentome combojiensis, Anentome spinosa,Anentome jullieni, Anentome scalarina and Anentome wykoffi. A.helena is widely distributed throughout Southeast Asia, including Cambodia, Indonesia, Malaysia and especially in northern Thailand[1]. A. helena is commonly known as the “assassin snail”and tend to live on soft, muddy bottoms[2-4]. While A. helena is popular in aquariums, its biology remains unclear. Thus, there is increasing apprehension about what would happen if this species was introduced outside its native range[5]. Moreover, A. helena in Chiang Mai province was infected with echinostome metacercariae[6].The parasites from the Family Echinostomatidae can infect the gastrointestinal tract of a variety of animals and humans, which leads to a disease called echinostomiasis. The disease has been generally found in north and northeastern parts of Thailand[7].Since the disease may be transmitted to humans by ingesting raw or partially cooked snails, there is substantial interest in understanding echinostome metacercariae infection in snail populations in order to control potential echinostomiasis cases in humans[8].
The taxonomic relationships among the members of Anentome are ambiguous. Traditional taxonomic classification of gastropods is commonly based on shell morphological characteristics, including shape, size and surface sculpture[1,9]. A review by Brandt[1]examined Thai freshwater snails and synonymised 7 species with Clea helena only based on shell morphology and noted that their shell was extremely variable with regard to size, shape and costulation. However, environmental factors can greatly affect A.helena shell morphology[10]. Thus, given the flourishing interest in this species across various disciplines, the taxonomy of this snail needs to be robustly explored.
Molecular phylogeny offers a framework to clarify organism classification[11]. During the previous decades, this technique has been used to resolve systematics and evolution issues when morphological and physiological characteristics prove ambiguous[12].Studies on freshwater snail phylogeny using molecular DNA sequences are potentially useful, and the most common genes utilized in these studies are mitochondrial cytochrome oxidase subunit I (COI) and 16S rRNA[13-16]. Ambiguities in the species emphasize the need for rigorous morphological analyses accompanied by molecular comparisons to clarify the species boundaries of A. helena and determine the parasitic infection of this snail. This research will provide useful information that may be valuable for developing public health prevention strategies for echinostomiasis and conservation management of A. helena in the future.
2. Materials and methods
2.1. Sampling and identification
A. helena s.l. samples were collected throughout northern Thailand by using the count per minute method (samples collected every 10 minutes per sampling by 5 collectors). Specimens were identified based on shell morphology and compared with original descriptions[1].
2.2. Shell and radula study
One hundred empty shells of each A. helena morphotype were investigated. Weight was determined using an electronic scale with 0.01 g precision. Shell height, shell width, aperture height and aperture width were measured with vernier calipers. In order to detect variations in shell morphology between the 2 morphotypes,the mean values of linear measurements (shell or aperture sizes)were compared by t-test (P<0.05) using IBM SPSS Statistics ver.22.0[17].
At least 30 radulae of each morphotype were examined. The radulae were extracted from the snail buccal cavity using a stereomicroscope, boiled in 5% sodium hydroxide for 5 min and then washed with distilled water. Dehydration was performed by immersing the radulae in increasing alcohol concentrations (10%,30%, 50%, 70%, 80% and 95%, respectively). Subsequently, the specimens were mounted on stubs with the help of a sharp-tipped needle on carbon conductive adhesive tapes. The stubs were then coated with gold and observed using a scanning electron microscope(JSM 5910 LV) at the Electron Microscope Research and Service Center, Faculty of Science, Chiang Mai University.
2.3. DNA extraction, PCR amplification and sequencing
Genomic DNA was extracted from foot tissues of 15 individuals using 150 μL of Chelex 100 and 3 μL of proteinase K. Samples were incubated at 55 ℃ for 1 h, 95 ℃ for 30 min and then stored at -20 ℃ until use. COI and 16S rRNA mitochondrial genes were amplified using the following primers: LCOI490 (5'-GGTCAACAAATCATAAAGATATTGG-3') and HCO2198(5'-TAAACTTCAGGGTGACCAAAAAATCA-3') for the COI gene[18] and 16sar (5'-CGCCTGTTTATCAAAAACAT-3') and 16sbr (5'-CCGGTCTGAACTCAGATCACGT-3') for the 16S rRNA gene[19]. PCR was performed in 20 μL reactions with 5 μL 2 Illustra Hot Start Master Mix (GE Healthcare), 1 M each primer,deionized water and approximately 10 ng DNA template. For COI,thermal cycling was: 94 ℃ for 2 min, 36 cycles of 94 ℃ for 30 s,42 ℃ for 2 min and 72 ℃ for 2 min, and a final extension step of 72 ℃ for 5 min. For 16S rRNA, thermal cycling was performed in the same way as for COI except the annealing temperature was changed to 49℃ for 30 s. The amplified products were checked with 1% (w/v) agarose gel electrophoresis using 0.5× TBE buffer. Gels were run at 100 V for 45 min and visualised with RedSafe nucleic acid staining solution and UV transillumination. PCR products were purified and sequenced using BigDye® Terminator v3.1 cycle sequencing kit chemistry and 1st BASE DNA Sequencing Services (Applied Biosystems).
2.4. Phylogenetic analysis
The taxonomic sampling scheme used herein included 17 individuals identified as A. helena sensu lato, 6 related species[16] and 6 specimens of Nassodonta dorri[3]. Chromatograms were visually inspected and edited as necessary with ClustalW[20] as implemented in MEGA 7.0[21]. COI was translated to check for amino acid sequence, stop codons and frameshift mutations. All newly generated sequences have been deposited in GenBank (Table 1). Phylogenetic trees were constructed using neighbour joining (NJ), maximum likelihood (ML) and Bayesian inference (BI). The phylogenetic trees were rooted on the genus Nassodonta since it has been suggested to represent close relatives of Anentome[3,16]. jModeltest 0.1.1[22]was used to evaluate the best evolutionary substitution model under Akaike Information Criterion[23]. NJ analysis was performed using PAUP*v4.0[24] based on appropriate models (16S: GTR+I+G; COI:GTR+G; concatenated dataset, Kimura 2-parameter (K2P)). One thousand replicates of bootstrap resampling[25] were performed to assign support to branches in the NJ tree. ML was performed using PhyML v.3[26], with randomised stepwise addition parsimony trees(number seed = 13) and with 1 000 bootstrap replicates[25,27]. BI analysis was performed using MrBayes v3.1.2[28], where the tree space was explored using 4 chains for 2 runs of a Markov chain Monte Carlo algorithm. The BI analysis was run for 7 000 000 generations with heating parameter set at 0.095, sampled every 100 generations, burnin set at 25%, and the last remaining trees were used to determine posterior probabilities of the BI tree. Convergence was observed by verifying that the average standard deviation of the split frequencies(between 2 runs) was below 0.01[29].
2.5. Parasitic infection
Infection of A. helena with trematode larva was examined by the crushing method. Metacercariae were covered with a cover slip and observed under a light microscope. The shape, size and cyst wall structure were used for characterisation of the metacercariae. The presence of collar spines was highly indicative of the echinostomemetacercaria type. Different numbers of arranged collar spines have been reported before raging from a minimum of 31 to a maximum of 51. The number of collar spines presented in each echinostome species can be used for identification[6]. So, the numbers of collar spines on the oral collar were counted[6,30]. The metacercaria were compressed, fixed in 5% formalin and stained with haematoxylin.They were dehydrated in ascending grades of alcohols, cleared in xylene and mounted in Permount for permanent slides. Two measures were calculated: prevalence of the parasite species as percentage of hosts infected by that species; intensity of the infection as the mean number of parasite larvae per infected host.
3. Results
3.1. Morphology
A total of 325 A. helena were collected throughout Thailand (Figure 1, Table 1). A. helena represents one of the most widespread species of Anentome. We collected the snail throughout northern Thailand.The A. helena shell was conical in shape and had dark brown and yellowish-tan bands around the whorls. The morphological examination of A. helena specimens revealed 2 distinct morphotypes.Morphotype A exhibited 2 elongated shell forms, while morphotype B exhibited globate shells (Figure 2A, C). As previously observed,A. helena were anatomically cryptic due to large shell structure variation. Average shell dimensions for morphotype A (Figure 2A)were 1.889 cm (range 1.600-2.300 cm) in height and 0.847 cm (range 0.700-1.000 cm) in width; average aperture sizes were 0.659 cm (range 0.590-0.720 cm) in height and 0.341 cm (range 0.300-0.380 cm) in width. For morphotype B (Figure 2C), average shell height was 1.996 cm (range 1.800-2.400 cm) and width was 0.928 cm (range 0.900-1.200 cm); aperture height was 0.753 cm (range 0.660-0.830 cm) and aperture width was 0.350 cm (range 0.30-0.44 cm). The shell and aperture widths were statistically significantly different (P<0.05)(Table 2).
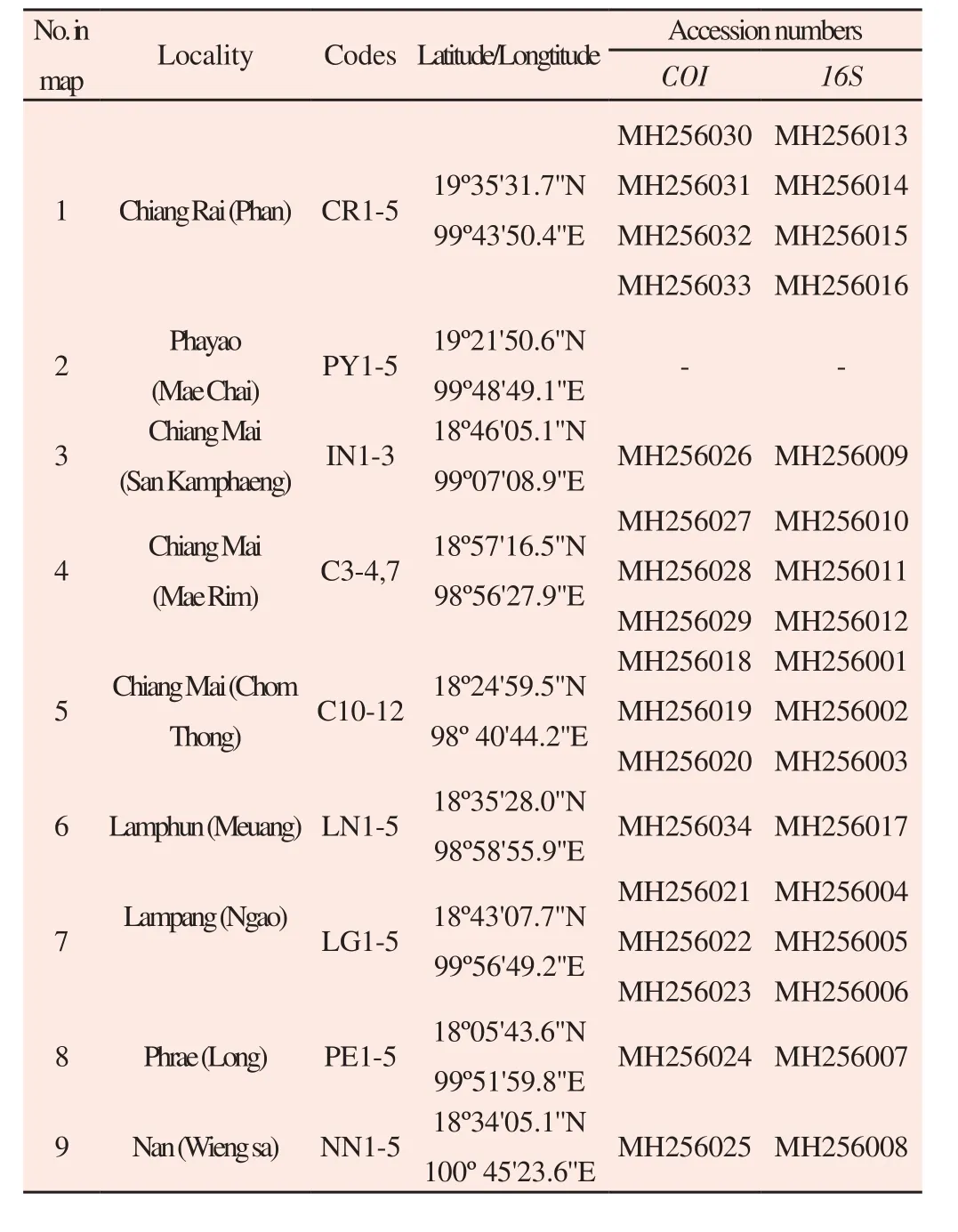
Table 1 List of localities and GenBank accession numbers of samples.
A. helena radulae were examined using a scanning electron microscope (350 magnification; Figure 2B, D). The radula was a stenoglossan type with a general formula of 1:1:1, each row of radula teeth consisted of 1 central tooth and 1 lateral tooth on each side. Examination of radular morphology also revealed differences between morphotypes (Figure 2B, D). Radulae of morphotype A(Figure 2B) consisted of approximately 6 cusps (range 5-8 cusps)of the central tooth and 3 cusps on each side of the lateral teeth.The secondary cusp of the lateral tooth was triangular in shape and narrower and shorter than the other cusps. Comparatively,morphotype B had a much more rounded cusp on the central part of the lateral tooth that was similartin shape to the other cusps, but with more convex bases compared to morphotype A. There were approximately 8 cusps (range 6-11 cusps) of the central tooth (Figure 2D). The outermost lateral teeth were similar between morphotypes.
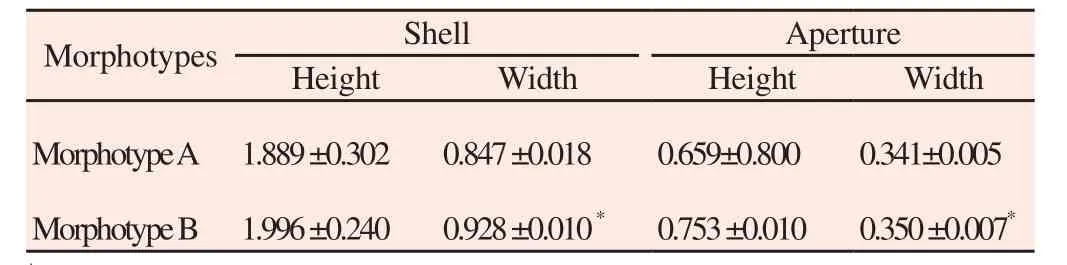
Table 2 Shell measurements of A. helena morphotypes.
3.2. Phylogenetic analyses
The COI and 16S rRNA-derived phylogenetic trees shared similar topologies that principally referred to relationships between the 2 morphotypes and division of the outgroup and other related species.All trees showed comparable evolutionary patterns with moderately high statistical support for each clade. The phylogenetic tree (Figure 3) was separated into 2 main clades: clade 1, all Anentome spp. and clade 2, outgroup (Nassodonta spp.). Clade 1 was also divided into 2 subclades: clade A, all Anentome spp. in Thailand and clade B,Anentome spp. from other countries; there was 100, 100 and 1.00 bootstrap support, respectively, from the NJ, ML and posterior probability of the BI method. Both morphotypes were placed in clade A and were further separated into clade A1 (morphotype A)and A2 (morphotype B) with moderately high statistical support and 61, 75 and 0.98 bootstrap support (NJ, ML and BI, respectively).

Figure 2. A. helena morphotype A and B.
3.3 Epidemiology
The prevalence of echinostome metacercariae in A. helena was examined from northern Thailand (Figure 4). Metacercariae type was identified by morphology. The 37 collar spines echinostome metacercariae were found and resembled members of Echinostoma group[6]. Infected snails were only recorded in San Kamphaeng district, Chiang Mai Province. The highest prevalence (7.5%) was found in A. helena. The highest intensity was noted in the same locality with (1.670±0.577) metacercariae per snail.

Figure 3. The ML tree obtained from sequence analysis of the concatenated COI and 16S rRNA dataset (1 178 bp).
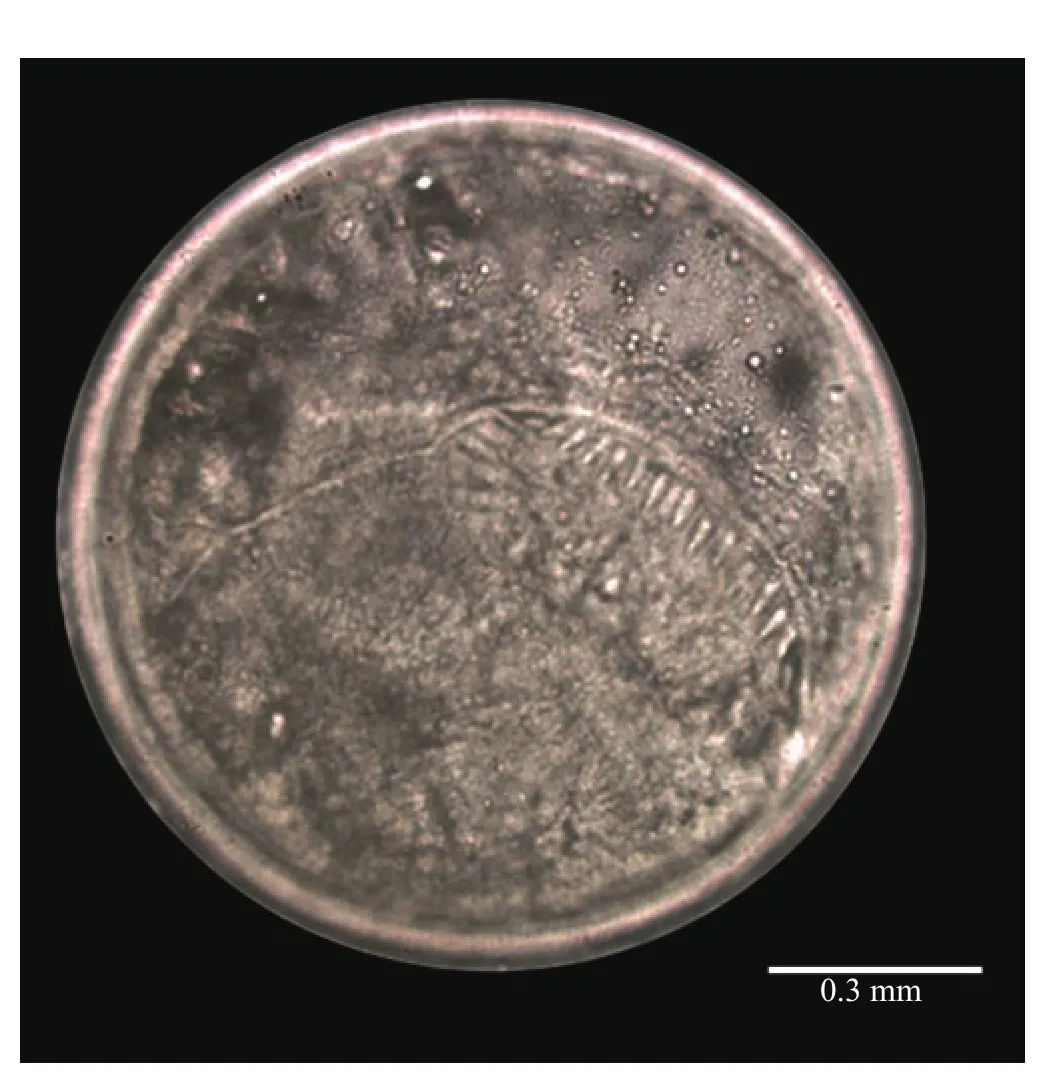
Figure 4. Echinostome metacercariae infected in A. helena from San Kamphaeng district, Chiang Mai Province.
4. Discussion
The molecular relationships among species of the genus Anentome are ambiguous[16]. Furthermore, the taxonomy of A. helena is unclear.Originally, this species had the combination name Melania helena (von dem Busch, 1847)[31], but subsequent alternative representation led to it being recognised as Clea (now Anentome) helena (von dem Busch,1847)[1] and also combined with many synonyms. Subsequently,Strong et al.[16] suggested that many taxa belonged to the genus Clea.Based on the morphology and molecular phylogeny evidence, they suggested that the species should be recognised as A. helena because of morphological differences in the shell, operculum and radula from Clea nigricans, the type species of Clea.
This study initially demonstrated cryptic diversity based on shell morphology in A. helena collected from different populations in northern Thailand. Phylogenetic analysis showed moderate support that A. helena morphotype A formed a sister group to morphotype B. The morphotypes exhibited different shell and aperture sizes as well as radular structure. Shell characteristics, including shape, size,colour pattern, sculpture and number of whorl have been used to identify the different Anentome species[1]. However, morphological classification based on a snail’s shell is occasionally misleading for accurate species delimitation[32,33]. In many cases, shell morphological variations may be caused by phenotypic plasticity in response to environmental conditions[32,33]. Our results showed significant differences between the two morphotypes that were supported by both morphological characteristics and phylogeny. The molecular results supported the recognition of two A. helena lineages from northern Thailand. A. helena morphotype A was recovered from sites in Chiang Mai (Chom Thong district) Lampang (Ngao district),Phrae (Long district) and Nan (Wieng Sa district; No. 9 on map),while A. helena morphotype B was recovered from sites in Chiang Rai (Phan district), Phayao (Mae Chai district), Chiang Mai (San Kamphaeng and Mae Rim districts) and Lamphun (Meuang district).This occurrence may be caused by allopatric speciation related to geographic isolation (most likely due to river systems in northern Thailand). Moreover, the current distribution of these populations may reflect the great drainage connectivity across the Chiang Mai and Lumpang river system[34] and/or may be accidental.
While both lineages received high support from analyses of the mitochondrial and concatenated datasets, specimens identified here as species from northern Thailand displayed high K2P average pairwise distances for COI and 16S (COI, 0.080;16S, 0.036-0.086) when compared with species from eastern Thailand and Malaysia . Thus, the geographic scale of this species complex, the morphological distinctiveness of the lineages and the molecular evidence are provided here. This study has uncovered an unrecognised radiation within A. helena from northern Thailand that was formerly recognised as a single species. Thus, comprehensive revision of snail systematics is needed. However, another complicating influence is that the freshwaters in Southeast Asia are affected by human impacts, including habitat loss, agriculture,pollution and impoundment, and many snail species are believed to already be extinct[35]. Nevertheless, the validity of the currently discovered species remains to be tested.
Our study provided epidemiological data of echinostome metacercariae in A. helena from northern Thailand between February 2017 and January 2018. Based on morphological characteristics,the worms showed a prominent head collar with 37 collar spines arranged in a double row. In this case, they were identified as genus Echinostoma. In this survey, we found different prevalence in echinostome metacercariae infection. Only the San Kamphaeng district, Chiang Mai Province contained A. helena infected with echinostome metacercariae; prevalence and intensity were 7.5%and 1.670±0.577 metacercariae per snail, respectively. The diverse ecosystem in the agricultural area of San Kamphaeng district may provide suitable ecological conditions that allow for the distribution of trematodes in the snails that differ from other localities[6]. The overall prevalence and intensity of metacercarial infection were 0.92% and 1.670±0.577, both of which are considered low infection rates when compared to other snail species (e.g., Filopaludina spp.and Lymnaea spp). These species were reported to act as the first and second most common intermediate hosts of trematodes in the family Echinostomatidae[6,8,36]. Parasite infection patterns are often related to host dispersal and gathering of parasites, principally during larval stages[37,38]. Moreover, reports have revealed an effect of parasite infection on the population structure, including death, of intermediate host snails[39]. Detection of this infection in this area is important for public health control with regard to monitoring dispersion; echinostomes can be easily transmitted to humans through eating raw or partially cooked snails. Coordinated parasite control strategies across the region are of great importance.
Conflict of interest statement
The authors declare that they have no competing interests.
杂志排行

Asian Pacific Journal of Tropical Medicine的其它文章
- Acute kidney injury in leptospirosis: Overview and perspectives
- In vitro antiproliferative and apoptosis inducing effect of a methanolic extract of Azadirachta indica oil on selected cancerous and noncancerous cell lines
- Efficacy of voriconazole on leishmaniasis by Leishmania major: An in vitro and in vivo study
- Anti-schistosomal activities of Echinops kebericho Mesfin root and Hagenia abyssinica (Bruce) J.F Gmel flower part crude extracts in Swiss albino mice
- Calcium carbonate supplementation causes memory impairment in mice
- In vitro anticancer activity of polysaccharide extracted from red alga Jania rubens against breast and colon cancer cell lines