Efficacy of voriconazole on leishmaniasis by Leishmania major: An in vitro and in vivo study
2018-11-14AhmadOryanSomayehBahramiEffatBemani
Ahmad Oryan, Somayeh Bahrami, Effat Bemani
1Department of Pathology, School of Veterinary Medicine, Shiraz University, Shiraz, Iran
2Department of Parasitology, Faculty of Veterinary Medicine, Shahid Chamran University of Ahvaz, Ahvaz, Iran
Keywords:Leishmania major Wound healing Azoles Voriconazole Histopathology
ABSTRACT Objective: To appraise the activity of voriconazole against Leishmania major (L. major) in vitro and its effectiveness on wound regeneration in cutaneous leishmaniasis in BALB/c mice.Methods: The IC50 of voriconazole against promastigotes and intra-macrophage amastigotes of L. major was investigated in vitro. The in vivo study was performed by treating the L. major infected BALB/c mice. When the wounds appeared in the base of tail, treatment was started by administration of 30 mg/kg voriconazole for 28 consecutive days orally. Results: The IC50 of voriconazole against promastigotes and intra-macrophage amastigotes were 0.74 and 0.89 μM, respectively. Voriconazole decreased lipid peroxidation and IL-6 level. Histopathological findings indicated accelerated healing in the voriconazole treated group compared to other groups. Conclusions: Our results demonstrate that voriconazole can be an option in treating the cutaneous leishmaniasis by L. major.
1. Introduction
Leishmaniasis is a vector-borne protozoan disease that includes both human and animals and is transmitted to hosts by female infected sand flies[1-3]. Its etiological agents, Leishmania spp., exist as flagellated extracellular promastigote in phlebotomine sand flies and aflagellated intracellular amastigote in mammalian hosts[2,4,5].The disease has multiple clinical forms; these clinical manifestations are affected by issues such as genetic diversity of parasite species,geographical origin of species, site of inoculation, infecting species,number of inoculated parasites, immune and nutritional status of host, and coexistence of immunosuppressive diseases[1,6-11].
Cutaneous leishmaniasis (CL) is the most usual clinical manifestation of leishmaniasis, and approximately thirty percent of cases happen in the Mediterranean basin, the Americas, and western Asia from the Middle East to Central Asia[1,12,13]. The geographic distribution and incidence of CL is increasing in recent years[2,3,6].On the other hand, an appropriate vaccine is not available to control the disease[9]. Therefore, hurried diagnosis and favored treatment of patients are necessary to control the disease[3]. Pentavalent antimonials like meglumine antimoniate and sodium stibogluconate are the first-line therapy for human leishmaniasis in many countries.Side effects, long-term treatment and increase in parasite resistance to drug limit usage of pentavalent antimonials. When treatment failure by pentavalent antimonials occurs, alternative drugs like miltefosine, pentamidine, paromomycin and amphotericin B, alone or in combination are prescribed. As these alternative drugs have limitations and side effects[1,5,8,9], therefore, many investigations have been performed to find a low-risk and more effective agent than the currently available drugs.
Usefulness of some azoles as promising drugs in treating leishmaniasis has previously been indicated in some in vitro[14,15],in vivo[16] and clinical studies[17-24]. Even, some cases of CL that did not respond to pentavalent antimonial were successfully treated with ketoconazole and itraconazole azoles[25,26]. Azoles are beneficial antifungal agents inhibiting cytochrome P-450-mediated 14-α-demethylation of lanosterol to block ergosterol synthesis in cellular membranes of eukaryotic microorganisms and subsequently promote membrane permeability[2,24,27,28]. Ergosterol is the main sterol needed for cell viability and growth of fungi and some of the Trypanosomatidae family, while it is absent in the host mammalian cells[29].
Voriconazole, approved in 2002, is a triazole with high bioavailability, large distribution volume and high affinity against yeasts, dermatophytes, and molds[27,30-32]. The drug is available as both intravenous and oral formulations and is currently used clinically for treatment of fungi infections[30,31]. The mechanism of action of voriconazole is to inhibit ergosterol biosynthesis, and disrupt membrane’s structure and function in fungi[31]. It has been approved that fluconazole is safe and effective for leishmaniasis therapy[20,24,33]. In other words, an in vitro investigation showed that the IC50of voriconazole against the promastigotes and axenic amastigotes of Leishmania spp. is superior to fluconazole[34].Therefore, we investigated the efficacy of voriconazole on the promastigotes and intra-macrophage amastigotes, intracellular form of the parasite, of Leishmania major (L. major). The action of voriconazole on wound healing, immune response, ferric reducing ability of plasma (FRAP), malondialdehyde (MDA) and adiponectin levels and antioxidant enzyme activities were also investigated in CL in BALB/c mice.
2. Materials and methods
2.1. Parasite and cell culture
The promastigotes of L. major (MRHO/IR/75/ER) were incubated at 26 ℃ in RPMI-1640 medium supplemented by 10% heatinactivated fetal calf serum (FCS), and 1% penicillin (50 U/mL)streptomycin (50 μg/mL) solution (Sigma, St. Louis, Mo., USA).Raw 264.7, a murine monocyte/macrophage cell line, was purchased from the Iranian biological resource center. The cells were cultured in RPMI-1640 plus 10% FCS and 1% penicillin (50 μg/mL)streptomycin (50 μg/mL) solution. The medium was then changed three times per week. After monolayer formation on the bottom subcultures were performed of the flasks.
2.2. Effects of voriconazole on L. major promastigotes
The L. major promastigotes were cultured in 96-well plates(1×106parasites/well) in presence of different concentrations of voriconazole purchased as Vfend. The wells with medium and promastigotes were used as positive control and the medium with no organism was considered as blank. Forty-eight hours after treatment,3-(4, 5-methylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (MTT)was added to cells in culture and the plate was incubated at 37 ℃for 4 h. Finally, DMSO was added to melt the formazan dye and absorbance was then measured at 570 nm. Percentage of viability was computed by: [(AT-AB)∕(AC-AB)]×100%.
Where, AB is OD of the blank well, AC is OD of the negative control and AT is OD of the treated cells.
2.3. Effects of voriconazole on macrophages
The cytotoxicity effects of voriconazole (0 to 50 μM) on raw 264.7 macrophage cell line was studied, using MTT assay as described by Lyu and Park[35]. The macrophages (106cells/well) were cultured in 96- well culture plates in absence and presence of voriconazole (0-50 μM) at 37 ℃ with 5% CO2for 24 h.
A total of 50 μL MTT was then added to each well. After four hours incubation with MTT, to dissolve the formazan dye, DMSO was added. Based on the absorbance of the treated and untreated specimen and the blank wells, the live cell counts were determined.The drug cytotoxicity was estimated as the dose that resulted in 50%mitigation in macrophage counts.
2.4. Effects of voriconazole on L. major amastigotes
Raw 264.7 macrophages were kept at 37 ℃ and 5% CO2in RPMI 1640 medium (Gibco, UK) plus 10% FCS. A proportion of 10 promastigotes per each macrophage was employed, for parasite infection, and the infected cells were incubated in 5% CO2at 37 ℃ for 24 h. The cultures were treated with various dilutions of voriconazole. The uninfected macrophages and the L. major-infected macrophages were respectively assessed as the negative and positive controls. The medium of culture chambers was removed, and the adhered macrophages were flashed by PBS. The macrophages were fixed and stained by Giemsa (1:10 dilution), and the multiplication index was calculated. Multiplication Index (MI) was estimated using the following formula: MI=(Mean number of amastigotes per 100 treated macrophages/Mean number of amastigotes per 100 control macrophages) ×100.
2.5. In vivo study
Female inbred, 5-7 weeks old BALB/c mice were purchased from Pasture Institute (Tehran, Iran). At beginning of the experiments, the mice were 6 to 8 weeks and weighted (19.11±1.12) g. The animals were kept in animal house of School of Veterinary Medicine, of our university, with free accessibility to food and water. Animal care and all experiments were performed as recommended by the Institutional Animal Care and Use committee of the School of Veterinary Medicine, Shiraz University, Shiraz, Iran, on 16 July 2016 (Ethical approval number 26-4-1395). Infectivity by a standard strain of L. major (MRHO/IR/75/ER) was maintained by way of regular passages in susceptible BALB/c mice at our Center of Comparative and Experimental Medicine. A total of 100 μL of phosphate-buffered saline containing 1×106amastigotes collected from the donor BALB/c mice were inoculated subcutaneously into the base of tails of mice. Five weeks after inoculation, the mice were randomly divided into four groups of six mice and treatment was started when a wound was present at the injection site. The animals were treated daily for 28 consecutive days with the drugs, as follows:group 1 without any treatment as the negative control group, group 2 was treated with Tween 80 (1%) used as vehicle orally, group 3 with glucantime at 60 mg/kg intraperitoneally, and group 4 with voriconazole at 30 mg/kg orally. Oral drugs were administered through a stomach tube.
The mice were euthanized after 28 days of treatment. The blood samples were transferred into sterile vacuum tubes containing EDTA for determination of antioxidant enzymes. The plasma was separated by centrifugation and stored at -70 ℃ until use to measure the MDA and adiponectin levels, and FRAP.
2.6. Measurement of the lesion size
Photographs were taken every four days since treatment began. The images were analyzed in a Digimizer 4.2.6.0 to compute the surface area of the wound. The mean and standard error were calculated for each experimental condition.
2.7. Smear preparation to estimate the parasite load
From the margins of lesions, Giemsa-stained smears were provided. To evaluate severity of infection, the mean amastigote counts in cytoplasm of fifty random macrophages was calculated.The infection intensity was estimated by scoring the infections based on the defined criteria[36,37], i.e., weak (<100 parasites/macrophage),moderate (100 -1 000 parasites/macrophage), and severe (>1 000 parasites/macrophage) infections in the macrophages.
2.8. Superoxide dismutase (SOD) and glutathione peroxidase(GPX) enzyme activity tests
A commercial kit (Ransod kit, Randox Com, UK) was used to measure the SOD activity. The value of the enzyme was calculated as units per gram of hemoglobin, while one unit of SOD corresponded to 50% inhibition of 2-(4-iodophenyl)-3-(4-nitrophenol)-5-phenyltetrazolium chloride (INT) reduction.
A Ransel kit, produced by Randox according to the method of Paglia and Valentine[38] was utilized to measure the GPX activity.Based on manufacturer’s instruction, the value of the enzyme was calculated as units per gram of hemoglobin.
2.9. Measurement of the FRAP
Based upon the Benzie and Strain[39] method, plasma antioxidant activity was evaluated using the FRAP-assay. In this method ferric tripyridyltriazine complex reduce to ferrous tripyridyltriazine at low pH and complex with blue color can be analyzed at 593 nm.
2.10. Measurement of adiponectin and MDA level
Adiponectin level of plasma was measured by enzyme linked immunoabsorbent assay kit (Bosterbio, China) based on recommended manual. The MDA level was measured as thiobarbituric acid-reactive material as recommended by Placer et al[40]. The method is relied on spectrophotometrical assay of color formation during the reaction to MDA with thiobarbituric acid. Plasma MDA level was calculated as the 548 nm absorbance opposed to bi-distilled water blank.
2.11. Preparing formalin-fixed Leishmania antigens
The formalin-fixed Leishmania antigens were prepared for in vitro lymphocyte proliferation and cytokine secretion assays. The L. major stationary promastigotes were collected and washed three folds in sterile PBS. The samples were fixed in 1% formal saline for 1 h and washed then three times in PBS. The parasites were counted in haemocytometer counting chamber and resuspended in a concentration of 5×108/mL in sterile PBS and kept at -70 ℃ until required.
2.12. Culture of lymph node cells and measurement of cytokines
After 4 weeks, the inguinal lymph nodes were excised and singlecell suspensions were obtained by teasing through a 70 μm mesh.Suspensions of single cell were cultured aseptically in RPMI-1640 medium (Shellmax, China) containing 2 mM L-glutamine, 100 U penicillin/mL, 100 μg streptomycin/mL, 10% heat inactivated FCS(Shellmax, China) and 0.05 mM beta-mercaptoethanol. The cells were distributed into 12-well plates at a concentration of 1×106cells/well and were cultivated with formalin- fixed antigens at 37 ℃in 5% CO2. The cell culture supernatants were harvested following 24 h and analyzed for tumor necrosis factor (TNF-α) or 72 h for interleukin-6 (IL-6). Levels of TNF-α and IL-6 were assessed by ELISA, using commercial kits (Bosterbio, China).
2.13. Histological analysis
To study the healing status of the cutaneous wounds, sections from the skin lesions were routinely prepared and stained by hematoxylin and eosin (H & E) and Masson’s trichrome.
2.14. Statistical analysis
In vitro anti-leishmanial activity, expressed as IC50(50% inhibitory concentration), was determined by linear regression analysis. Oneway analysis of variance (ANOVA) was used for the statistical analysis of differences between the groups by SPSS version 16.0. A P-value of fewer than 0.05 was regarded to represent a significant difference.
3. Results
3.1. In vitro studies
The cytotoxicity concentration of voriconazole to reduce 50%of viable macrophages was 3 μM. Viability of the L. major promastigotes and intra-macrophage amastigotes in presence of voriconazole or glucantime was evaluated. The half inhibitory concentration was estimated 0.74 and 0.89 μM for promastigotesand intra-macrophage amastigotes incubated with voriconazole for 48 h, respectively and 0.25 and 0.40 μM for promastigotes and intra-macrophage amastigotes incubated with glucantime for 48 h,respectively.
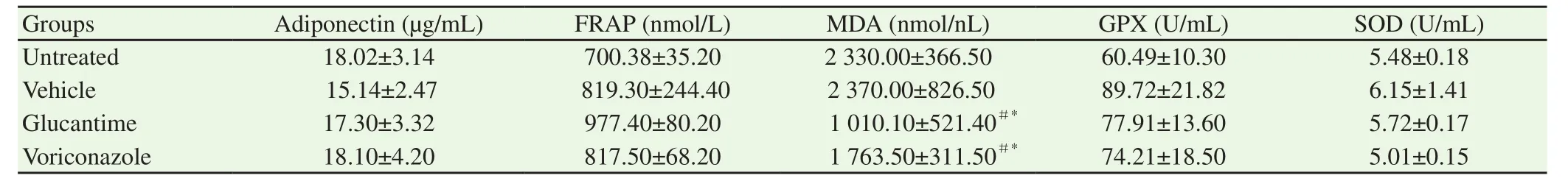
Table 1 Antioxidant enzymes activities and MDA, FRAP and adiponectin concentration in plasma of the treated and controls groups.
3.2. In vivo studies
3.2.1. Size of the lesions
The mean area of lesions, measured every four days at beginning of treatment, diminished in the mice treated with voriconazole and glucantime compared to the mice treated with vehicle and untreated mice (Figure 1), but this downgrading was not statistically significant.

Figure 1. Changes in the surface area, in mm2 of the treated and untreated lesions every four days at the beginning of treatment in the L. major infected BALB/c mice for 28 days.
3.2.2. Scoring of the parasite load
Parasite intensity in the margin of lesions decreased in the glucantime and voriconazole groups in comparson to the vehicle and untreated groups. The mean amastigote counts in cytoplasms of fifty random macrophages obtained weak (<100 parasites/macrophage)for glucantime group, moderate (100-1 000 parasites/macrophage)for voriconazole group and severe (>1 000 parasites/macrophage)for vehicle and untreated groups.
3.2.3. Antioxidant enzyme activities and plasma level of FRAP, adiponectin and MDA
As demonstrated in Table 1, the MDA level reduced significantly in the voriconazole and glucantime treated groups compared to the untreated groups and vehicle (P<0.05). There were no significant differences between the four groups in the level of GPX and SOD activity and also plasma level of FRAP and adiponectin (P>0.05).
3.2.4. Level of TNF-α and IL-6
As indicated in Table 2, the level of IL-6 in supernatants of the inguinal lymph node cell culture was significantly lower in the voriconazole and glucantime treated groups (P<0.05). The level of TNF-α was significantly lower in the glucantime group compared to the voriconazole, vehicle and untreated groups (P<0.05).

Table 2Level of TNF-α and IL-6 in cell culture supernatants of inguinal lymph node of the treated and un-treated groups (pg/mL).
3.2.5. Histological analysis
There were numerous inflammatory cells, amastigotes (as free or intracellular) and extensive necrosis in tissue sections of the lesions of vehicle and untreated groups (Figure 2). Intensity of necrosis and inflammation diminished in the voriconazole and glucantime treated groups. As shown in Table 3, statistically, reduction of neutrophils and macrophages, and increment of fibroblast and fibrocyte counts(granulation tissue-forming cells) was significant in the voriconazole and glucantime treated groups (P<0.05). The fibroblast and fibrocyte counts and number of the newly formed blood vessels was significantly higher in the mice treated with voriconazole (30 mg/kg/day) compared to those treated with glucantime (60 mg/kg/day).As demonstrated in Figure 3, collagen deposition in the voriconazole and glucantime treated groups was greater than the vehicle and untreated ones.

Table 3 Number of cells in histopathological sections.
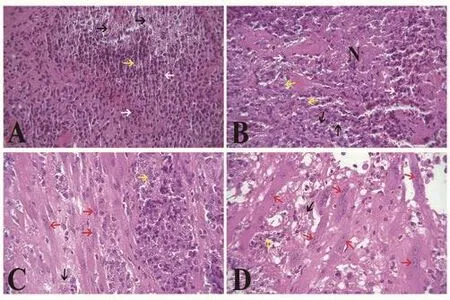
Figure 2. Histological sections of skin in the L. major infected BALB/c.Treated by voriconazole (D) for 28 days, compare to the glucantime (C) and vehicle (B) treated and the untreated (A) groups (H & E, ×400). Necrotic area (N) and number of macrophages (yellow arrow), neutrophils (white arrow) and amastigotes (black arrow) decreased in the voriconazole and glucantime groups. Presence of fibroblasts (red arrow) and neovascularization in depth of ulcers in the mice treated by voriconazole and glucantime indicated improved granulation tissue formation in the lesions of these therapeutic regimens.

Figure 3. Masson’s trichrome staining of lesions of the L. major infected BALB/c mice.Treated with voriconazole (D) for 28 days in comparison to the glucantime(C), vehicle (B) and untreated (A) groups (×400). Collagen deposition(arrow) enhanced in the voriconazole and glucantime treated groups in comparison to the untreated and vehicle groups.
4. Discussion
In vitro effects of some azoles against the Trypanosomatidae family has been approved in different studies[14,41,42]. The IC50of posaconazole against Trypanosoma cruzi epimastigote and amastigote forms obtained 25.0 nM and 1.0 nM, respectively[42]. Modification in the mitochondrion-kinetoplast region, formation of cytoplasmic autophagic vacuoles, formation of vesicles in the flagellar membrane and plasma membrane shedding were seen in transmission electron microscopy ultramicrographs of epimastigotes and intracellular amastigotes of Trypanosoma cruzi treated with posaconazole[42].It has previously been indicated that ketoconazole can induce a dose-dependent reduction in Leishmania infantum promastigotes and proliferation of axenic amastigotes[14]. Antiproliferative,physiological and ultrastructural studies performed by de Macedo-Silva et al[41] indicated that posaconazole and itraconazole are potent inhibitors of the Leishmania amazonensis growth. The IC50against promastigotes and intra-macrophage amastigotes of Leishmania amazonensis was 2.74 and 1.63 μM, respectively for posaconazole and it was 0.44 and 0.08 μM for itraconazole, respectively. These two drugs lead to alternations in mitochondrial function and structure by collapsing the mitochondrial membrane potential and swelling and rupturing the membrane, and additionally accumulation of lipid bodies and autophagosomes[41].
In our investigation, the IC50of voriconazole, a second-generation azole, was calculated 0.74 and 0.89 μM for L. major promastigotes and intra-macrophage amastigotes, respectively. This result is in agreement with the report by Kulkarni et al[34] that showed the in vitro effectiveness of voriconazole against leishmanial species.Difference in IC50value for L. major promastigotes estimated in our study and Kulkarni et al[34] [(2.130±0.176) μg/mL] may be due to presence of genetic dissimilarity in L. major that likely influence on sensitivity and response of parasite to the same drug[1,2,10,28].
It seems voriconazole (30 mg/kg/day) has more potency in accelerating wound healing than glucantime (60 mg/kg/day) in the same course of treatment (28 day) because the mean number of neovascularization, and fibroblast and fibrocyte counts in the mice treated with voriconazole was significantly higher compared to the mice treated with glucantime. However, voriconazole indicated lower intrinsic activity against L. major promastigotes and intra-macrophage amastigotes than glucantime. Also, consistent with in vitro results, in vivo study demonstrated glucantime and voriconazole can significantly reduce the parasite intensity in margin of lesion; however the mean counted amastigote aggregation inside macrophages was fewer in the glucantime treated group than the voriconazole treated ones. Although, TNF-α level was lower in the voriconazole treated group but this differences were not significant with the untreated and vehicle groups. However, decrease in the TNF-α level was significant in the glucantime treated group than untreated, vehicle and voriconazole groups. Baltch et al[43]claimed that voriconazole, itraconazole, and fluconazole effects on production of TNF-α by the human monocyte-derived macrophages was negligible. But Silva et al[32] suggested that the level of TNF-α in the lung homogenates of mice with cryptococcosis treated by voriconazole depended on the drug concentration administered.It has been indicated that TNF-α in combination with interferongamma (IFN-γ) activate macrophages to kill Leishmania through nitric oxide production[44,45]. Injection of murine recombinant TNF-α in the lesion of infected mice lead to significant reduce lesion development[46]. Therefore, TNF-α, a cytokine with both pro- and anti-inflammatory properties, is an essential component in L. major infections which contribute in control of the inflammatory reactions and parasite killing[44]. Any other way, its excess level causes damaging effects to the host’s cells[47].
Although, CL, in most cases, is a nonfatal and self-healing disease but complete cure can take months to years. Additionally, secondary infection, functional impairment, development of disfiguring permanent scars, parasite dissemination to mucous membranes or relapse may happen through recovery period[1,2,7,8,17]. Therefore,the main purposes in CL therapy are to reduce the recovery period and prevent the parasite propagation by improving the host’s ability to heal the lesions and to eradicate the microorganism[2].Our histopathological results indicated that voriconazole can accelerate wound healing of CL caused by L. major by improvement of granulation tissue formation. The mechanism of action of voriconazole is possibly through reducing the oxidative stress and IL-6 levels[48]. IL-6 is a glycosylated protein with 184 amino acids. This proinflammatory cytokine is secreted by neutrophils,monocytes, macrophages, fibroblasts, mast cells, smooth muscle cells, endothelial cells, T-cells, B cells and dendritic cells throughout inflammation or infection[49,50]. IL-6 is an essential cytokine in the primary phase of wound healing[51] but its high level in chronic wounds implies to persistent inflammation and leads to impaired wound healing[52,53]. In addition, IL-6 contributes to acute stage response by inducing acute phase response protein synthesizing in hepatocytes, and by transmission of acute inflammation to chronic one by monocyte recruitment to inflammatory area[54],differentiating B cells to antibody secreting plasma cells by triggering IL-21 production in CD4+T cells[49,55], promotion of T helper (Th2) differentiation by activating CD4+T to IL-4 producing and inhibiting Th1 differentiation by interfering with IFN-γ signaling[56]. Activation of Th1 cells is suitable for efficient clearance of an intracellular agent such as Leishmania spp. and recovery of disease by producing IFN-γ and TNF-α while Th2 cell response is preferred in extracellular infections by activating B cells and antibody production and can prevent progression of the disease caused by extracellular microorganisms[56].
Adiponectin can increase type I collagen production by dermal fibroblasts and promote keratinocyte proliferation and migration[57,58]. Therefore, adiponectin is a dominant mediator which is responsible to control cutaneous wound healing[58]. In the present study, no significant difference was seen in adiponectin level between the groups. However, the MDA level, the main product to evaluate lipid peroxidation[59,60], was significantly lower in the voriconazole treated mice. The principal mechanism in Leishmania eradication is oxidative burst using reactive oxygen species (ROS)and nitric oxide[61]. Conversely, ROS have unpaired electrons in their structure and react with nucleic acids, carbohydrates, lipids and proteins in cell[59] and its overproduction or impaired detoxification of oxidants causes oxidative stress[59,62] leading to cellular damage and impaired wound repair and subsequently development of chronic, non-healing wounds[62-64]. Therefore, regulation of ROS production and detoxification is a pivotal factor for a normal wound repair process. Both non-enzymatic antioxidants and enzymatic antioxidants detoxify ROS[64]. The level of non-enzymatic antioxidants measured by FRAP and the efficacy of antioxidative enzymes (SOD, GPX), in the present study, was not significantly different between the treated and un-treated groups.
In summary, our study recommended that voriconazole is effective against promastigote and amastigote of L. major and can enhance healing of the CL in BALB/c mice probably by modulating the IL-6 level and reducing oxidative stress. Therefore, voriconazole may be a suitable drug option in treatment of CL. However, the antileishmanial effects of voriconazole remains incomplete at present and is an area for future research to characterize the mechanisms of activity and the effect of voriconazole in humans with cutaneous leishmaniasis.
Conflicts of interest statement
The authors declare that there is no conflict of interest.
杂志排行

Asian Pacific Journal of Tropical Medicine的其它文章
- Acute kidney injury in leptospirosis: Overview and perspectives
- In vitro antiproliferative and apoptosis inducing effect of a methanolic extract of Azadirachta indica oil on selected cancerous and noncancerous cell lines
- Anti-schistosomal activities of Echinops kebericho Mesfin root and Hagenia abyssinica (Bruce) J.F Gmel flower part crude extracts in Swiss albino mice
- Calcium carbonate supplementation causes memory impairment in mice
- In vitro anticancer activity of polysaccharide extracted from red alga Jania rubens against breast and colon cancer cell lines
- Investigation of cryptic diversity and occurrence of echinostome metacercariae infection in Anentome helena (von dem Busch, 1847)