自噬在孤独症模型大鼠行为学及海马PSD-95蛋白表达变化中的作用
2018-11-07罗瑜平陈运华童雪涛
刘 芬,周 波,文 敏,罗瑜平,陈运华,艾 戎,童雪涛
孤独症,又称自闭症,是一种神经发育障碍性疾病,主要表现为行为重复刻板和社会沟通障碍,部分患者伴有智力障碍,其发病率约0.6%,男女发病比率约为4 ∶1,迄今为止,其发病机制尚不明确[1-2]。近年来,遗传学和基因组学研究鉴定的大量孤独症相关候选基因,如NLNG3/4、NRXN1、TSC1/2、FMR1等,其中大部分基因与突触发育有关[3-6],突触已成为孤独症研究领域的焦点。
近年来,越来越多的证据表明,突触的发育与自噬密切相关,自噬在突触发育中的作用已经在果蝇和线虫模型中得到证实,但其在哺乳动物突触发育中的作用仍不清楚[7]。本实验通过在Wistar鼠孕12.5 d给于腹腔注射丙戊酸诱导孤独症动物模型,运用自梳理实验及三箱实验验证模型是否成功,并给予自噬增强剂雷帕霉素(rapamycin)和自噬抑制剂(3-methyladenine)干预孤独症大鼠,再次运用自梳理实验及三箱实验检测自噬干预前后孤独症大鼠行为变化,Western blot方法对比自噬干预前后孤独症大鼠海马自噬相关蛋白(LC3-Ⅱ、Beclin 1)与突触相关蛋白(PSD-95)的表达变化,探讨自噬的变化在孤独症大鼠突触发育中的可能机制。
1 材料与方法
1.1材料健康繁殖期 Wistar雌鼠20只(体质量250~260 g)和雄鼠10只(体质量270~280 g),贵州医科大学实验动物中心提供 (合格证号SCXK京[2014-0004]),丙戊酸钠 (sodium valproate,VPA )(P4543)和LC3-Ⅱ兔抗鼠多克隆抗体(L8918)购自美国Sigma公司;雷帕霉素(Rapamycin)(R-5000)购自上海睿铂赛生物科技有限公司;3-methyladenine(3-MA)(HY-19312)购自MCE公司;BCA蛋白质定量试剂盒、RIPA裂解液、Beclin 1兔抗鼠多克隆抗体(bs-1353R)、β-actin兔抗鼠多克隆抗体(bs-0061R)购自北京博奥森生物技术有限公司;PSD-95兔抗鼠多克隆抗体(#3409)购自美国CST公司。
1.2方法
1.2.1模型建立 参考Schneider et al[8]建模方法,选取健康成年Wistar雌鼠20只和雄鼠10只,雌雄鼠按2 ∶1比例于19:00合笼过夜,用苦味酸给实验动物做标记编号,次日早晨进行雌鼠阴道涂片,光镜观察,涂片上观察到精子的当天计为妊娠(embryonic day,E)第1天 (E1),孕鼠分笼饲养。在E12.5 d 时,实验组孕鼠给予250 g/L的VPA (600 mg/kg)腹腔注射,产下的子代鼠为孤独症模型组,对照组孕鼠腹腔注射同等剂量的生理盐水,产下的子代鼠为正常组。所有子代鼠出生当日计为生后第1天 (postnatal,P1)。于P35在孤独症模型组中随机取30只分为三组:自噬增强组给予腹腔注射雷帕霉素(5 mg/kg,溶于二甲基亚枫和PBS)1周;自噬抑制组给予腹腔注射3-MA(5 mg/kg,溶于PBS)1周,模型组给予同等剂量的溶剂腹腔注射。
1.2.2自梳理实验 自梳理实验用于描述大鼠刻板、重复行为。P42,将各组大鼠分别放置于标准鼠盒内(长30 cm×宽25 cm×高20 cm ),盒底覆盖1 cm敷料,以阻止大鼠挖掘盒底,待大鼠在盒内适应5 min后,开始视频记录10 min,观察每只大鼠的活动行为,再用秒表计数10 min内每只大鼠用于梳理身体各区域的累计时间。
1.2.3三箱实验 三箱实验用于检测大鼠社会交互行为和好奇行为。自制木箱(长120 cm×宽45 cm×高50 cm),木箱中间用2块木板隔开,使中间格长60 cm,左右格长均为30 cm,2块隔板下方各有1个大小适宜且可开关的门,可允许大鼠通过,在2边格子中各放2个铁丝鼠笼(可容纳1只大鼠)。P42,将待测大鼠放置中间格子中,并关闭两侧可控门让大鼠适应5 min后,打开两侧可控门,使大鼠在三个格子中可自由通行,再适应5 min,然后在左边格子的鼠笼中放置 1只陌生鼠(stranger1),视频记录10 min,随后在右边格子的鼠笼中放置另一只陌生鼠(stranger2),且stranger1不取出,视频记录10 min,分别计数2个10 min内大鼠在左右格子中的活动时间。
1.2.4Western blot法检测LC3-Ⅱ、Beclin 1和PSD-95的表达 于P42冰上断颈处死大鼠,迅速取大鼠海马,称重后放入EP管中,加入适量RIPA裂解液(使用前已加入苯甲基磺酰氟PMSF)提取蛋白,用BCA蛋白定量试剂盒测定总蛋白浓度,取40 μg总蛋白样品于10%SDS-PAGE分离,半干转方法将蛋白转到聚偏二氟乙烯(PVDF)膜上,于5%脱脂牛奶封闭1 h,加入相应抗体(LC3-Ⅱ兔抗鼠多克隆抗体1 ∶1 000、Beclin 1兔抗鼠多克隆抗体1 ∶1 000、PSD-95兔抗鼠多克隆抗体1 ∶1 000、β-actin兔抗鼠多克隆抗体1 ∶10 000)4 ℃孵育过夜,次日洗膜后加入相应二抗(山羊抗兔IgG 1 ∶10 000)室温孵育2 h,洗膜,ECL发光液显影曝光,用Image J软件对条带进行半定量分析。
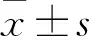
2 结果
2.1行为学检测自梳理实验结果显示:大鼠自梳理身体各区域时间,模型组(45.9±13.97)比正常组(25.4±9.00)明显增加(F=3.842,P<0.01),与模型组(治疗后)(47.2±13.93)比较,Rap组梳理时间(28.3±11.26)显著减少,3-MA组梳理时间(71.3±8.84)显著增加(F=34.898,P<0.01),提示模型组有重复行为,而Rap组重复行为较模型组有所减轻,3-MA组重复行为较模型组进一步加重。
三箱实验结果显示:前10 min,模型组在放有stranger1的格子中停留时间较正常组明显减少(P<0.01),而在放有空鼠笼(empty)的格子中停留时间较正常组明显增加(P<0.01);与模型组(治疗后)比较,Rap组在stranger1的格子中停留时间增加(P<0.05),在empty的格子中停留时间减少(P<0.05);3-MA组则相反,在stranger1的格子中停留时间减少(P<0.05),而在empty的格子中停留时长差异无统计学意义(P>0.05),3-MA组社会交往能力较模型组减弱。后10 min,模型组在放有stranger1的格子中停留时间较正常组增加(P<0.05),而在放有stranger2的格子中停留时间较正常组减少(P<0.05);与模型组(治疗后)比较,Rap组在stranger1的格子中停留时间明显减少(P<0.01),在stranger2的格子中停留时间明显增加(P<0.05);3-MA组在stranger1的格子中停留时间增加(P<0.01),在stranger2的格子中停留时间减少(P<0.01)。见表1。
2.2Westernblot检测LC3-Ⅱ、Beclin1和PSD-95的蛋白表达水平与正常组比较,模型组海马LC3-Ⅱ、Beclin 1蛋白表达水平下调(P<0.05),PSD-95蛋白表达水平上调(P<0.05);与模型组(治疗后)相比,Rap组LC3-Ⅱ、Beclin 1蛋白表达水平上调(均P<0.05),PSD-95蛋白表达水平下调(P<0.05),3-MA组LC3-Ⅱ、Beclin 1蛋白表达水平下调(均P<0.05),PSD-95蛋白表达水平上调(P<0.05)。见图1、2。
3 讨论
3.1VPA化学干预法可成功诱导孤独症动物模型VPA是一种广泛使用的药物,主要用于预防癫痫、偏头痛和双相情感障碍[7, 9],在母孕期暴露于丙戊酸可引起神经系统损伤。人类胚胎的中枢神经系统发育始于神经管,胚龄20~24 d为神经管闭合时间,这段时间对应到大鼠E12.5期间,该时间窗对孕鼠进行VPA治疗可诱导孤独症样行为[9-10]。近年来,众多证据表明,母孕期暴露于丙戊酸可增加自闭症谱系障碍(ASD)的患病风险[11]。VPA诱导的子代鼠出现社会交往障碍[12],重复和刻板行为[8, 12],这些发现为孤独症模型的建立提供有力的证据。本研究参考Schneider et al[8]建模方法,该模型的解剖及发病机制与人类的十分相似[13],该动物模型中可以观察到VPA大鼠行为改变类似于孤独症患者行为障碍。自梳理实验显示,模型组较正常组理毛时间明显延长,提示模型组有重复、刻板行为;三箱实验显示,模型组在放有stranger1的格子中停留时间较正常组减少,而在放有空鼠笼的格子中停留时间较正常组增加,提示模型组缺乏社会交往能力,且模型组在放有stranger1的格子中停留时间较正常组增加,而在放有stranger2的格子中停留时间较正常组减少(P<0.05),提示模型组缺乏对新鲜事物的偏好,孤独症患者主要表现为重复、刻板行为及社会交往障碍。本研究方法所诱导的孤独症行为成功的模拟了孤独症患者的行为障碍,表明该模型建立成功。
3.2增强自噬促进孤独症模型大鼠突触发育突触是神经元之间在功能上发生联系的部位,也是信息传递的关键部位,其数量及结构的完整性对维持脑功能的正常发挥起着重要作用。突触发育依赖于突触相关蛋白合成与降解之间的动态平衡[14]。Mashayekhi et al[15]提出SHANK3作为孤独症发病机制的重要候选基因,该基因的缺失将导致突触功能障碍;相关研究[3, 16]显示mTORC1-eIF4E 通路异常激活,mRNA过度翻译,导致突触相关蛋白合成异常增强,突触发育失衡,引起孤独症。可见,突触相关蛋白合成与降解失衡,导致突触发育障碍,可能是引起孤独症的重要机制之一[7]。本课题组在前期研究中也证实孤独症大鼠前额叶皮质 synaptophysin、PSD-95 等突触相关蛋白表达异常增高。增强对过剩的突触相关蛋白的降解,恢复合成与降解间的动态平衡成为孤独症治疗的关键策略之一。自噬溶酶体系统是体内重要的蛋白降解途径,通过去除受损的细胞器和降解长寿或易聚集蛋白对细胞内环境平衡起着重要作用[4-5,7],增强自噬可能通过促进突触蛋白的降解,改善孤独症的社交行为障碍。本研究通过母孕期化学干预法建立孤独症模型,证实孤独症大鼠自噬活性降低,突触相关蛋白PSD-95异常增高,进一步运用自噬增强剂(雷帕霉素)和自噬抑制剂(3-MA)于P35干预孤独症模型,结果显示增强自噬促进了突触相关蛋白PSD-95的降解,显著改善了社会交互障碍和刻板重复运动等孤独症样行为;反之,给于自噬抑制剂(3-MA)干预后,突触相关蛋白PSD-95进一步增多,孤独症样行为进一步加重。可见,增强自噬可改善孤独症大鼠突触发育, 进一步改善孤独症大鼠行为,这一结果与Sato et al[6]研究一致,其研究发现运用自噬增强剂雷帕霉素处理孤独症、结节性硬化症小鼠模型,发现增强自噬可显著逆转小鼠模型的社会交互障碍。Tang et al[7]研究发现mTOR过度活化,导致自噬缺陷,引起孤独症样行为改变,并引起突触功能障碍,雷帕霉素治疗可改善孤独症样行为和突触发育。
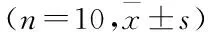
表1 各组大鼠三箱实验比较
与正常组比较:*P<0.05,**P<0.01;与模型组(治疗后)比较:#P<0.05,##P<0.01
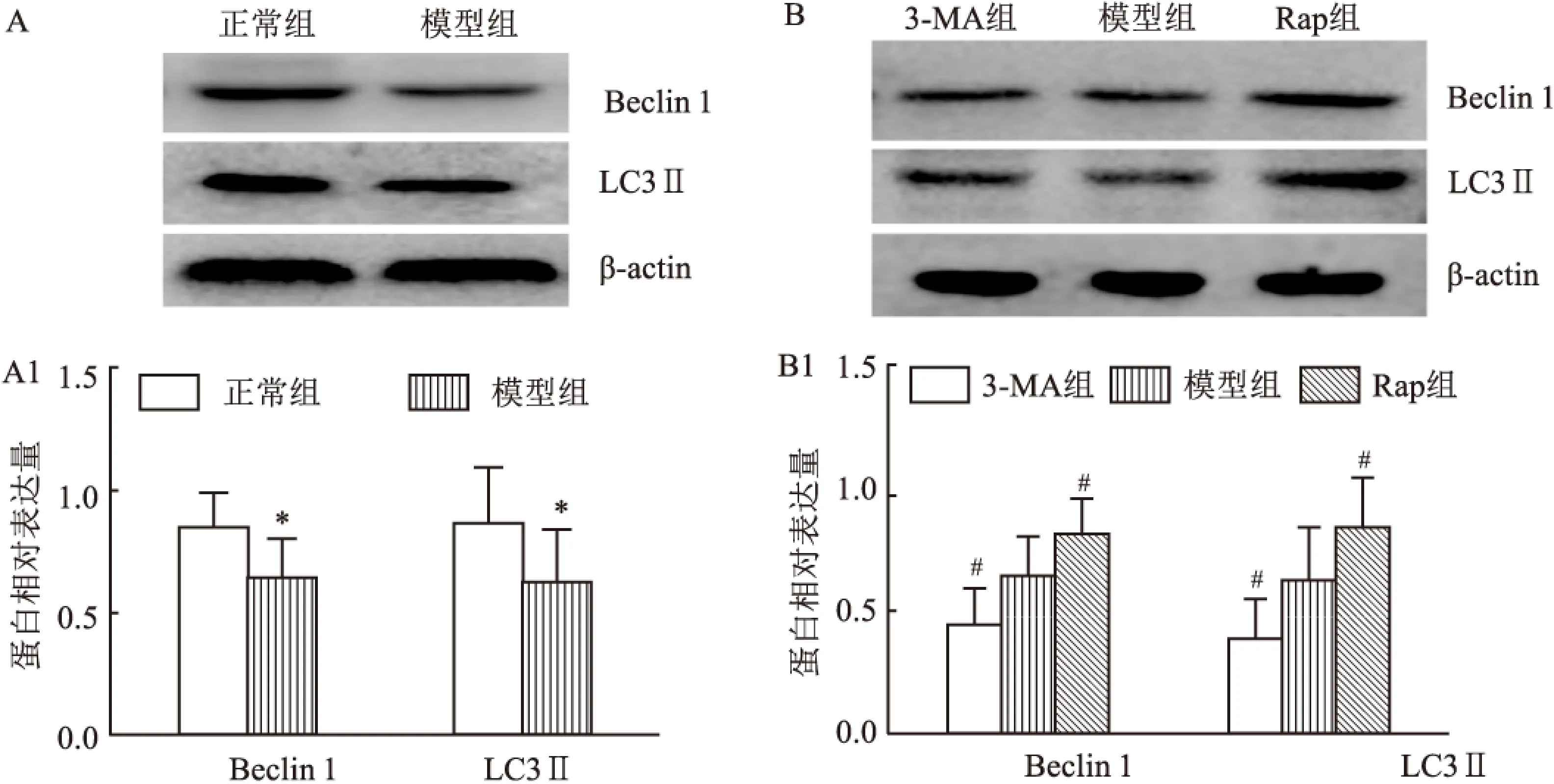
图1 各组海马Beclin 1、LC3 Ⅱ蛋白表达比较
A:正常组与模型组Beclin 1、LC3Ⅱ免疫印迹检测结果;A1:正常组与模型组Beclin 1、LC3Ⅱ蛋白表达量比较;B:模型组(治疗后)与3-MA组和Rap组Beclin 1、LC3-Ⅱ免疫印迹检测结果;B1:模型组(治疗后)与3-MA组和Rap组Beclin 1、LC3Ⅱ蛋白表达量比较;与正常组比较:*P<0.05;与模型组(治疗后)比较:#P<0.05

图2 各组海马PSD-95蛋白表达比较
A:正常组与模型组PSD-95免疫印迹检测结果;A1:正常组与模型组PSD-95蛋白表达量比较;B:模型组(治疗后)与3-MA组和Rap组PSD-95免疫印迹检测结果;B1:模型组(治疗后)与3-MA组和Rap组PSD-95蛋白表达量比较;与正常组比较:*P<0.05;与模型组(治疗后)比较:#P<0.05
