核苷酸转移酶与玉米非生物胁迫响应
2018-09-27陆佳运宋剑波王晓彦莫小为王碎抗许欣彤劳康文莫蓓莘
陆佳运,宋剑波,2,王晓彦,莫小为,王碎抗,许欣彤,劳康文,莫蓓莘
1)深圳大学生命与海洋科学学院,广东省植物表观遗传学重点实验室,广东深圳518060;2)江西农业大学理学院,江西南昌330045
核苷酸转移酶(nucleotidyl transferase protein,NTP) 蛋白是一类包含有poly(A) 聚合酶和末端核甘酸转移酶结构域的蛋白,在真核生物中核苷酸转移酶可以催化核糖核酸(ribonucleic acid,RNA)的非模版加尾,从而增强或减弱RNA的稳定性[1-3].根据底物的不同,核苷酸转移酶可分为尿苷酸转移酶、腺苷酸转移酶、胞苷酸转移酶和胸腺苷酸转移酶.核苷酸转移酶在人类[4]、酵母[5]、藻类[6]、果蝇[7]和线虫中[8]均有研究.植物核苷酸转移酶最早在拟南芥(Arabidopisisthaliana,At)中被发现[9-10],拟南芥中HESO1(HEN1 SUPPRESSOR1)能够催化非甲基化的miRNA 3′端非模板地添加尿苷酸,从而促进miRNA的降解.随后,拟南芥中与HESO1有协同作用的另一个核苷酸转移酶URT1(UTP:RNA URIDYLTRANSFERASE1)也被发现,URT1能对非甲基化的miRNA以及mRNA 3′端非模板地添加尿苷酸,从而影响RNA的稳定性[11-14].除了HESO1和URT1外,拟南芥中还有8个NTP相关基因,其功能还有待研究[12].核苷酸转移酶在不同的植物中是很保守的,说明它们可能具有重要的生物学功能.
植物的生长依赖于光、温度、水分、空气和有机营养物质.由于植物不可移动的特点,外界环境的胁迫会影响植物生长发育.各种胁迫对植物的影响往往同时发生,植物细胞内一系列的基因调控和变化具有“cross talk”现象[15].如干旱胁迫会使植物矮小、根系减少[16 ];植物激素脱落酸(abscisic acid,ABA)与冷、干旱、盐、重金属胁迫有明显关系[17].对水稻(Oryzasativa,Os)的研究发现,水稻中的NTP蛋白在非生物胁迫条件下被诱导表达[18],说明水稻的NTP基因可能参与了植物抗逆过程.玉米(Zeamays,Zm)是世界主要粮食作物之一,研究NTP相关基因与玉米植株抗逆之间的关系对提高玉米对逆境胁迫的耐受性及现代化育种具有重要指导意义.
1 材料与方法
1.1 材料及玉米NTP相关数据来源
利用Hmmer 3.0 (http://hmmer.janelia.org/)寻找玉米蛋白组中含有核苷酸转移酶结构域(PF01909) 的NTP蛋白. 核苷酸转移酶结构域(PF01909)来源于 Pfam 数据库(http://pfam.xfam.org).玉米和拟南芥基因以及蛋白序列信息分别从玉米数据库网站(https://www.maizegdb.org/)以及拟南芥信息资源网站(http: //www.arabidopsis.org/)下载.水稻蛋白信息来源于植物基因组学网站(https:/ /phytozome.jgi.doe.gov/pz/portal.html), 玉米B73种子为本课题保存.
1.2 玉米NTP蛋白的序列分析结果分析
利用MEGA5.0软件neighborjoining法则的P距离(P-distance)模型构建进化树,Bootstrap method值为1 000.利用Plantcare软件(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)分析启动子上游顺式作用胁迫元件.
1.3 非生物胁迫下NTP基因的表达分析
玉米幼苗在霍格兰德营养液中培养两周后分别进行:① 高盐处理.将幼苗转移到含300 mmol/L氯化钠的霍格兰德营养液中,分别处理0、3、6和12 h,两株幼苗为1个生物学重复,收取3个生物学重复.② 干旱处理.将幼苗转移至滤纸上,分别处理0、3、6和12 h,两株幼苗为1个生物学重复,收取3个生物学重复.③ ABA处理.将浓度为100 μmol/L的ABA溶液均匀喷洒在幼苗叶片上,分别处理0、3、6和12 h,两株幼苗为1个生物学重复,收取3个生物学重复.
利用Trizol法提取玉米RNA,反转录成cDNA,设计24种NTP基因的反转录聚合酶链式反应 (realtime fluorescence quantitative polymerase chain reaction, RT-PCR)引物(表1)分析处理不同时间后样品中NTP基因表达量的变化.
2 结果与分析
2.1 植物NTP基因的进化树分析
通过构建系统进化树对拟南芥、水稻和玉米的NTP蛋白进行分析(图1),在以上植物中都发现了NTP蛋白.拟南芥有10种NTP蛋白,其中,关于HESO1(AtNTP1)与URT1(AtNTP3)的研究较多.水稻与拟南芥NTP蛋白的种类数目较为接近,水稻有13种NTP蛋白,而玉米有24种NTP蛋白.通过进化树分析,发现这3类植物中的NTP蛋白具有高度的同源性.例如,ZmNTP24、OsNTP3与拟南芥中已知的NTPAtHESO1在同一分支上,说明同源性很高,ZmNTP16、OsNTP10与AtURT1具有高度同源性.进一步分析发现,玉米与水稻的NTP基因同源性更高,如ZmNTP7、ZmNTP11与OsNTP9,ZmNTP21与OsNTP4,ZmNTP1与OsNTP11、OsNTP12等.
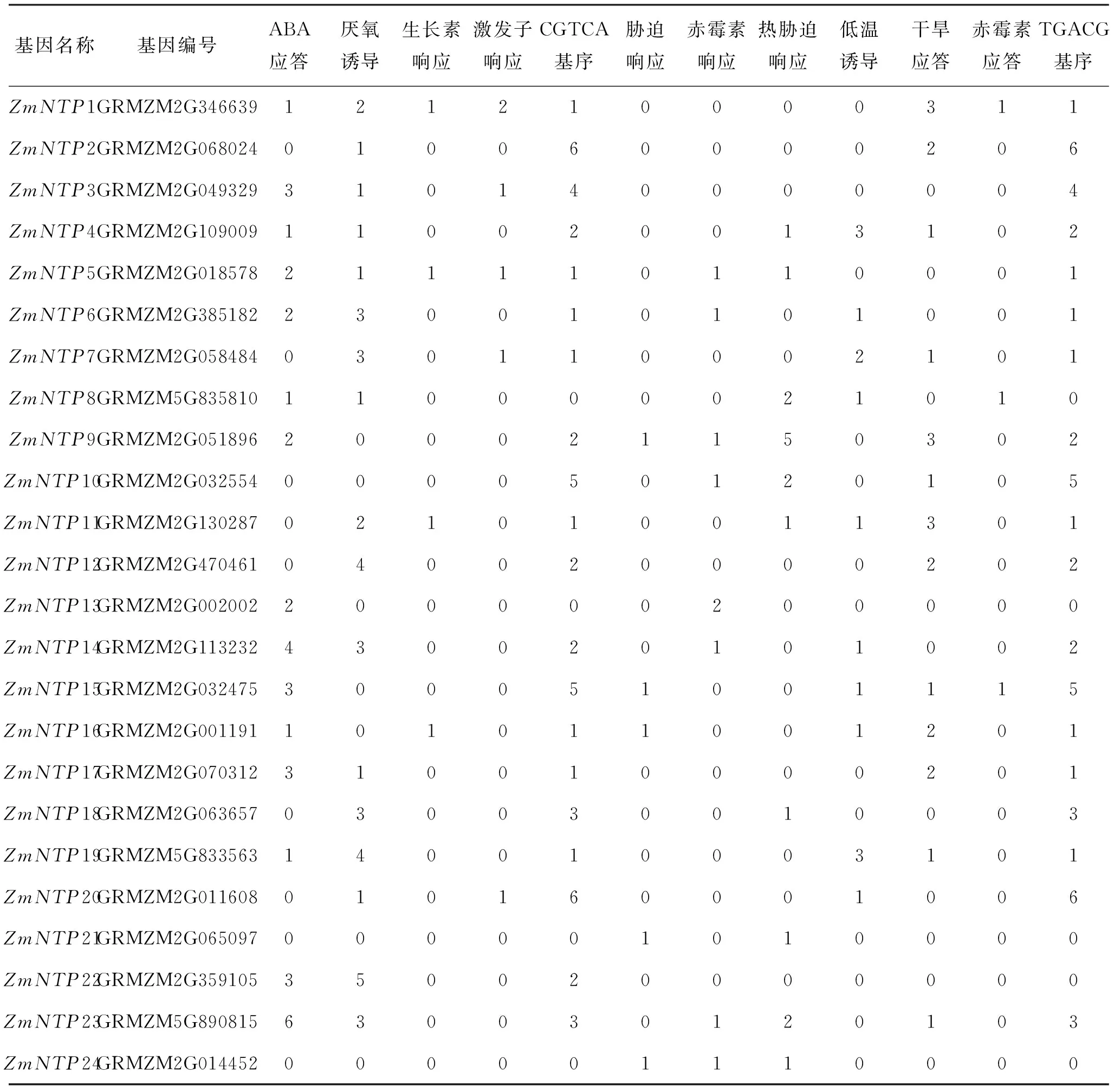
表1 玉米NTP基因胁迫相关的启动子元件分析
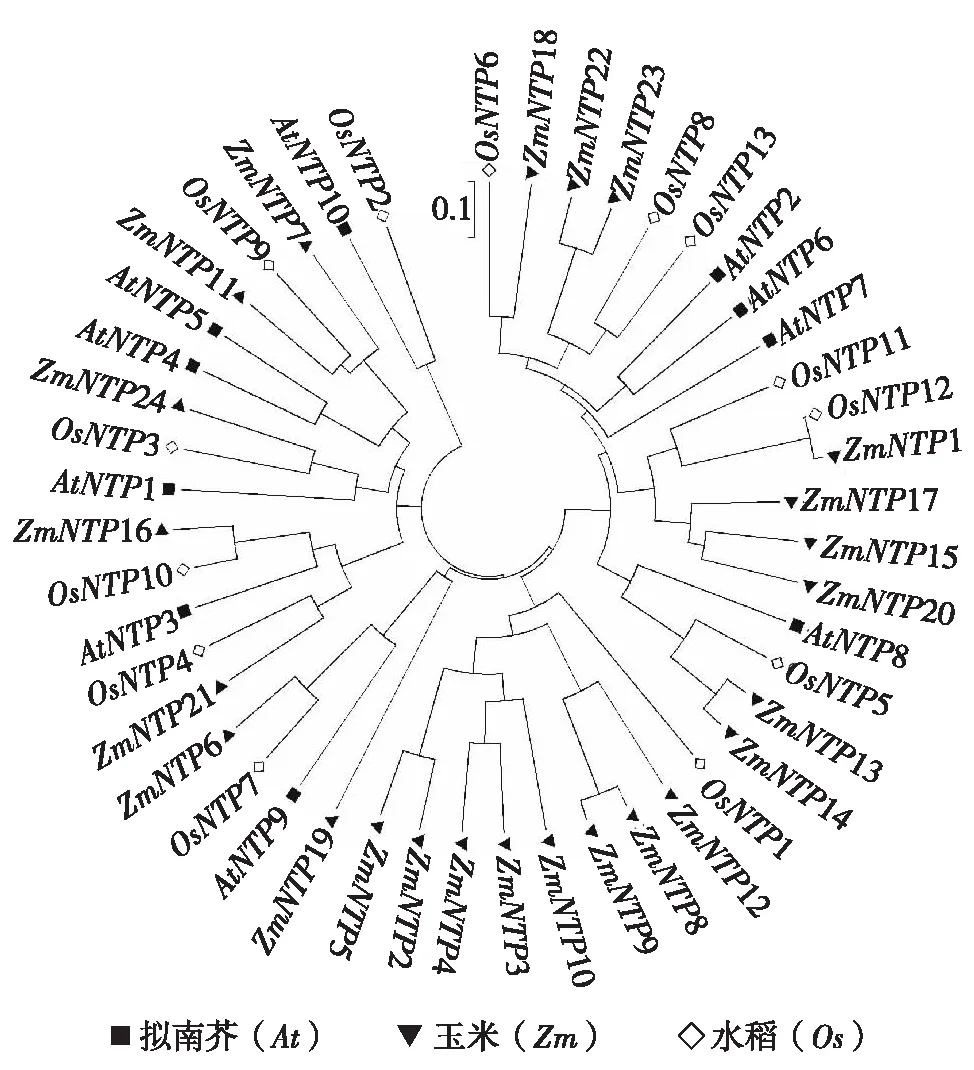
图1 NTP进化树Fig.1 Phylogenetic tree of NTP
2.2 玉米中NTP基因的启动子元件分析
为了研究玉米NTP基因与非生物胁迫之间的关系,通过生物信息分析软件Plant CARE分析了玉米NTP基因转录起始位点上游1 500碱基对的启动子序列.结果显示,所有NTP基因都具有一个或者多个非生物响应的相关元件(表1).ZmNTP3、ZmNTP14、ZmNTP15、ZmNTP17、ZmNTP22和ZmNTP23含有3个或以上的ABA响应元件; 24个NTP基因中有14个含有干旱应答元件;绝大多数的NTP基因都含有厌氧诱导元件.以上结果说明,核苷酸转移酶相关基因可能参与植物的非生物胁迫响应.
2.3 NTP基因在盐、干旱及植物激素ABA处理下的表达量分析
为验证玉米中核苷酸转移酶相关基因启动子元件分析的结果,对生长14 d的玉米幼苗进行了干旱和高盐(300 mmol/L)两种非生物胁迫处理.此外,因为ABA通路是植物应对外界环境变化的重要通路之一,所以用ABA(100 μmol/L)溶液喷洒玉米幼苗叶片进行胁迫处理.玉米的根分为胚根与节根,且根系相当发达,根组织与地上部分组织应对非生物胁迫的机制可能存在差异,所以将材料分为地上部分组织与根组织分别进行研究.玉米NTP基因的RT-PCR分析结果说明,在干旱、高盐和ABA处理下,核苷酸转移酶相关基因的表达量发生了显著的变化,部分核苷酸转移酶相关基因能被诱导表达(图2和图3).
ZmNTP22含有3个ABA顺式作用元件,在根组织中,ZmNTP22在ABA处理6 h后,表达量明显增多,在处理12 h后表达量达到最大,是对照的3.5倍左右(图3).值得注意的是,不同于根组织表达量的显著增加,ZmNTP22在地上部分的表达水平只有对照组的1/10(图2),这说明ZmNTP22在不同的组织中应答ABA胁迫的机制不同. 另外,ABA处理后,地上部分组织中(图2),ZmNTP2的表达量随着处理时间而上升;ZmNTP19在ABA处理3 h后表达量达到最大,随后下降.
ZmNTP9、ZmNTP17和ZmNTP23含有一个或以上的干旱应答元件,RT-PCR分析结果显示,受到干旱胁迫时,其表达量有显著提升.ZmNTP9有3个干旱应答元件,在受到干旱胁迫3 h后,其在根组织的表达量提升约3倍,随后降至对照组水平(图3).ZmNTP17含有两个干旱应答元件,在受到干旱胁迫3 h后,其在根组织的表达量提高了4倍随后恢复到对照水平(图3).ZmNTP23含有1个干旱应答元件,在受到干旱胁迫3 h后,在地上部分组织中表达量上升了3.5倍(图2);根组织中,ZmNTP23表达量对比对照组,在3、6和12 h时均有显著提升(图3).还有一些启动子不含干旱应答元件的NTP基因也明显受干旱胁迫的诱导表达.例如,干旱胁迫后,ZmNTP24无论在地上部分或是根组织中的表达量均有显著提升.在对比地上部分与根NTP基因表达差异时,发现在地上部分组织面对干旱胁迫时,ZmNTP1、ZmNTP5、ZmNTP6、ZmNTP9、ZmNTP10、ZmNTP11、ZmNTP12、ZmNTP16、ZmNTP17、ZmNTP18、ZmNTP19、ZmNTP20、ZmNTP21和ZmNTP22的表达量相比对照组均显著降低,表达量明显升高的仅有ZmNTP23和ZmNTP24;而在根组织中,受到干旱胁迫诱导表达升高的NTP基因总数达到10种(ZmNTP2、ZmNTP3、ZmNTP4、ZmNTP8、ZmNTP9、ZmNTP17、ZmNTP20、ZmNTP22、ZmNTP23和ZmNTP24). 说明玉米面对干旱胁迫时,其NTP基因可能主要在根组织中表达而非地上部分.
盐胁迫是植物生长环境中主要威胁之一.在受到盐胁迫后,地上部分组织中所有NTPs的表达量变化不大(图2).对比之下,在根组织中,ZmNTP4、ZmNTP8、ZmNTP9和ZmNTP22表达量有显著提升(图3);ZmNTP4和ZmNTP22在受到盐胁迫处理12 h后表达量升至1.5倍;在处理3 h时,ZmNTP8上升至1.5倍,ZmNTP9上升至3.5倍.此外,盐胁迫下,根组织中ZmNTP2、ZmNTP3、ZmNTP7、ZmNTP11、ZmNTP13、ZmNTP15、ZmNTP19和ZmNTP21的表达量均有显著下降(图3),而地上部分组织中仅有ZmNTP10和ZmNTP21表达量有明显降低(图2).总的来说,在面对盐胁迫时,地上部分仅有两个NTP基因的表达发生明显变化,而根组织中却有12个NTP基因出现显著性变化.这可能是因为玉米NTP基因面对不同非生物胁迫时有不同的响应机制,面对盐胁迫时NTP基因主要是通过根组织发挥作用.

图2 非生物胁迫下玉米地上部分组织中NTP基因表达分析Fig.2 The relative expression of ZmNTPs-shoot under abiotic stress
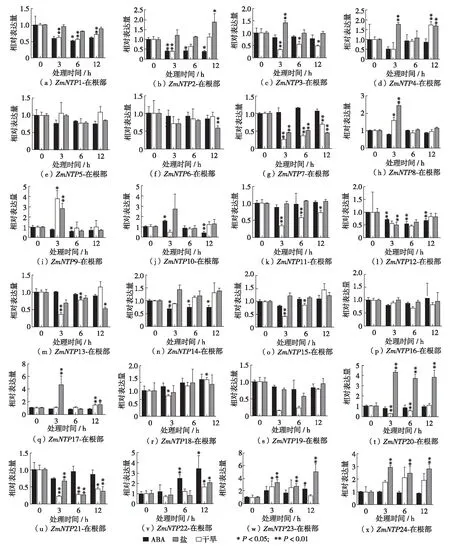
图3 在非生物胁迫下玉米根组织中NTP基因表达分析Fig.3 The relative expression of ZmNTPs-root under abiotic stress
3 讨 论
通过对玉米中24种NTP基因分析研究,生物信息学分析结果表明,存在于拟南芥、水稻与玉米中的NTP蛋白具有高度的保守性与同源性.拟南芥中含10种NTP蛋白,水稻中含有13种,玉米中多达24种NTP蛋白,这可能是物种基因组的复杂与冗余程度递增所致.植物中存在许多与胁迫相关基因,能通过改变这些基因的表达调控自身对于胁迫的耐受性. RT-PCR分析结果显示玉米中NTP基因能够在胁迫下诱导表达,表明这些NTP蛋白参与了植物抵抗外界胁迫的途径.这些NTP蛋白很有可能是通过对一些胁迫相关的RNA 3′端进行修饰,从而改变植株对胁迫的抵抗能力.这些关键的RNA可能是mRNA、小RNA或是长链非编码RNA.在对比玉米NTP蛋白在地上部分与根组织中表达差异时,结果表明,在面对不同类型胁迫时NTP基因表达有组织特异性,且在面对同一种胁迫时,NTP基因在地上部分组织与根组织中的表达调控机制也是不同的.虽然研究表明玉米NTP基因可能参与了抵抗非生物胁迫的相关途径,但相应调控的分子机制尚待进一步研究.
