水分胁迫对冬小麦冠层辐射截获率和利用效率的影响
2018-09-17李华龙窦子荷蒋腾聪何建强
李华龙 窦子荷 蒋腾聪 冯 浩 于 强 何建强
(1.西北农林科技大学旱区农业水土工程教育部重点实验室, 陕西杨凌 712100; 2.西北农林科技大学中国旱区节水农业研究院, 陕西杨凌 712100; 3.中国科学院水利部水土保持研究所黄土高原土壤侵蚀与旱地农业国家重点实验室, 陕西杨凌 712100)
0 引言
大田条件下,作物产量与冠层截获入射辐射能力以及辐射转化为干物质的效率密切相关[1]。冠层光合有效辐射(Photosynthetically active radiation,PAR)截获率是评价冠层辐射截获能力的有效指标,其中叶面积指数(Leaf area index,LAI)是影响冠层PAR截获能力最重要的因子[2]。作物通过叶片将到达冠层的光合有效辐射截获并通过光合作用转化为干物质[3],这个过程的转化效率被定义为辐射利用效率(Radiation use efficiency,RUE)。作物的干物质生产和冠层光截获率与RUE密切相关。GALLAGHER等[4]第一次确定了谷物类作物RUE的本质,即单位截获的太阳辐射所能产生干物质的量。MONTEITH等[5]将地上部累积生物量和累积辐射截获量线性回归模型的斜率定义为作物的RUE,因此地上部生物量可以通过累积辐射截获量与作物RUE相乘来计算。RITCHIE等[6]和SINCLAIR等[7]将这种关系应用到包括小麦、玉米等作物生长模型中,对作物产量进行估算。关于冠层PAR截获率和RUE的研究更多集中于群体结构、灌溉制度、种植方式等因素对其的影响。陈雨海等[8]研究发现,适当提高种植密度能够增加冠层辐射截获率,但截获率过高反而导致RUE降低。李全起等[9]、杨国敏等[10]研究表明,灌溉和秸秆覆盖有利于冬小麦生育后期冠层上部PAR截获率和RUE的提高。此外,合理的作物种植行距能够有效改善冠层结构分布和提高作物冠层消光系数,进而提高冠层光截获率和RUE,这在王兴亚等[11]、刘铁东等[12]的研究中得到充分体现。此外,许多研究表明环境胁迫(养分、水分)能降低作物叶片光合速率,减少作物生物量,进而减小作物的RUE[13-16]。DRECCER等[17]发现氮素胁迫能够影响小麦的生长并且降低其开花前的RUE。RODRGUEZ等[18]研究发现,小麦在出苗后的60 d里,RUE由于磷元素亏缺而出现减小的现象。水分胁迫会严重影响小麦的叶片生长,改变小麦LAI的垂直分布,使小麦在生长过程中的PAR截获量更少,进而影响光合同化物分配。HUSSAIN等[19]的研究也发现在巴勒斯坦地区干旱胁迫对小麦的RUE产生了负面效应。此外,RUE因水分胁迫而减少的现象在豌豆、花生[20-21]等作物中也被观测到。
冠层PAR截获率和RUE是作物生长模型、NPP遥感测定模型的重要参数之一。在众多模型中干物质形成是基于逐日冠层PAR截获率和RUE的函数,冠层PAR截获率的准确计算取决于模型对LAI和消光系数的充分描述[6]。RUE是模型中重要的输入参数,在模型中RUE是以不同的常数形式作为生态型参数给出的。但是在不同的气候条件和种植管理措施下,作物的RUE往往有很大差异。O’CONNELL等[20]报道的在澳大利亚半干旱区小麦的RUE是1.81 g/MJ,而JAMIESON等[22]在新西兰的研究得到的小麦RUE取2.2 g/MJ。此外,并没有发现针对冬小麦不同生育阶段的RUE变化规律的研究。RUE作为经验性参数的本质以及较低的估算精度都会使模型模拟结果产生极大的不确定性[23]。因此,研究冬小麦不同生育期受旱条件下的RUE能够帮助作物模型获得合理的参数数值,进而为提高作物模型的产量预测精度提供帮助。
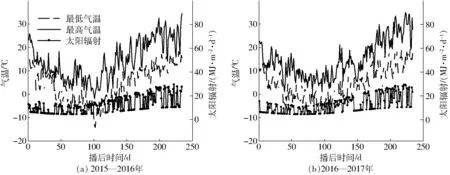
图1 2015—2016年和2016—2017年两季冬小麦生育期内逐日气温和逐日太阳辐射Fig.1 Daily air temperature and solar radiation during two wheat growing seasons of 2015—2016 and 2016—2017
小麦是中国的主要粮食作物,截止2017年,中国小麦的总产和播种面积分别占粮食作物的20.9%和21.4%。关中地区是我国主要的冬小麦种植区之一,该地区年降雨量600~700 mm,降水时空分布不均匀,其中5—10月的降水约占全年降水量的79.3%,冬春干旱加剧了冬小麦生长季的干旱程度[13]。探究水分胁迫条件下冬小麦冠层PAR截获率和RUE的变化规律及范围,能够更加深入理解水分胁迫对冬小麦生长发育和产量形成的影响机制。当前对于冬小麦冠层截获率的研究大多集中于冬小麦抽穗期以后,而关于冬小麦RUE的研究更是集中于整个生育期平均状况,并没有研究针对冬小麦各个生育期的RUE进行系统分析。此外,在冠层截获辐射测量方面,现有的研究普遍采用手持式的线性量子传感器定时定点测取[24-27],测量的时间间隔较长,测量时刻也多集中在正午,这可能导致约10%的冠层辐射截获率估算误差[28-29]。针对这些问题,为了降低辐射截获率计算误差,本研究采用安装在田间的PAR传感器全天候、连续监测冬小麦冠层顶部和底部的辐射通量。对关中地区冬小麦不同生育期受旱处理条件下的冠层辐射进行连续2年的观测研究,以揭示不同受旱条件下冬小麦冠层光截获率的变化规律;研究水分胁迫对RUE的影响以及各生育期RUE的变化范围,从而为作物模型生态型参数RUE的准确确定提供参考。
1 材料和方法
1.1 试验区概况
田间试验于2015年10月—2017年6月在陕西杨凌西北农林科技大学节水灌溉试验站(34°17′ N,108°04′ E,海拔506 m)进行,共种植2季冬小麦。试验站所在地区位于关中平原旱作区,属于暖温带季风气候区,年内降雨分布不均,冬小麦生育期内降水较少。试验区土壤为塿土,1 m深土层土壤pH值为8.14,有机碳含量为8.20 g/kg,全氮含量为0.62 g/kg。
在2015—2016生长季冬小麦抽穗前后的日平均太阳辐射强度分别为10.42、19.44 MJ/(m2·d),而在2016—2017生长季冬小麦抽穗前后日平均太阳辐射强度分别为10.22、20.85 MJ/(m2·d),此外,2年冬小麦生育期内累积太阳辐射分别为2 880.86、2 914.05 MJ;在气温方面,2015—2016年小麦抽穗前后的日平均气温分别为7.1、19.2℃,而2016—2017生长季小麦抽穗前后日平均气温分别为7.3、20.6℃;冬小麦2个生长季内生育期平均气温分别为9.2、9.7℃。整体而言,2016—2017生长季比2015—2016生长季有更多的辐射和积温(图1)。
1.2 田间试验设计
本试验供试冬小麦品种为关中地区主栽品种之一的“小偃22”,播种日期分别为2015年10月15日和2016年10月16日,播种方式为条播,播种深度5~6 cm,种植行距25 cm,种植密度400万株/hm2。为了保障小麦能够均匀出苗,各试验小区在播种前7 d灌水100 mm。各处理均在播前施用140 kg/hm2N和50 kg/hm2P2O5作为底肥,生育期内不再追肥。试验小区规格为4 m×2 m,各小区之间铺设1.5 m深的聚乙烯塑料隔离层以防止侧渗。试验区上方建有活动遮雨棚,降水时关闭以杜绝降水对试验结果的影响。
将冬小麦整个生育期划分为越冬期、返青期、拔节期、抽穗期和灌浆期等5个不同生长阶段,每相邻2个生长阶段连续受旱。根据冬小麦各个生育期的需水情况分别设置4个不同的处理:生育前期的返青+拔节受旱(Early stress, ES)、生育后期的抽穗+灌浆受旱(Later stress, LS)、全生育期不灌水(Whole stress, WS),以及每个生长阶段充分灌水处理(CK)作为对照。每个处理设置3个重复,在遮雨棚下按完全随机区组试验设计布设小区(表1)。陕西关中地区冬小麦全生育期需水量为400~500 mm,本研究选择400 mm作为CK处理的灌溉定额,试验分5次灌溉,灌水定额为80 mm,灌水方式为畦灌,采用水表计量灌水量,2年的试验处理相同。
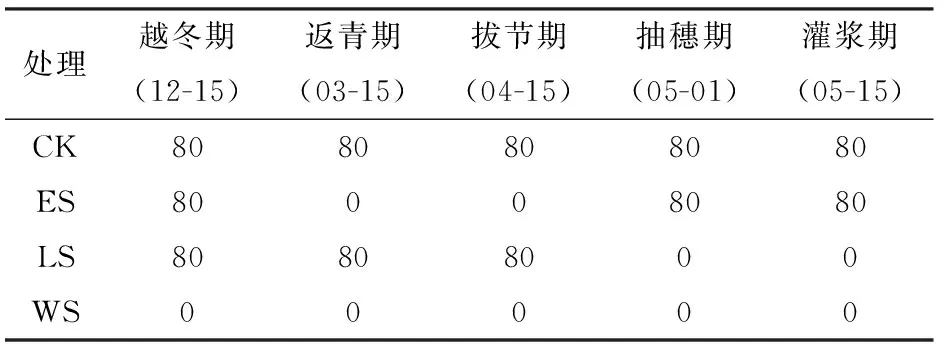
表1 不同生长阶段冬小麦受旱试验灌水处理Tab.1 Treatments of field experiment about winter wheat growth under water stress at different development stages mm
1.3 观测指标与方法
1.3.1土壤水分及小麦长势
土壤含水率采用烘干法测定。在小区内选取植株覆盖代表性样点,利用土钻每20 cm取一次样,取样深度为1 m,将土样装入铝盒混合均匀,放入烘箱在105℃下烘干至恒重。冬小麦拔节前每14 d取样一次,拔节后每7 d取样一次。冬小麦的物候期按照ZADOKS等[30]的方法每7 d观测一次,当各小区50%冬小麦进入了某一生育期标志该小区冬小麦达到相应生育期。
冬小麦出苗后在各试验小区选取能代表小区整体长势的1 m2植株样方,用于最终产量测定。全生育期内地上部生物量每14 d测一次,每次随机选取3株代表性植株,首先记录样品的分蘖数,随后在105℃下杀青30 min,75℃下干燥直到恒重后称量得到冬小麦干物质量。叶面积指数(Leaf area index,LAI)采用Sunscan 冠层分析系统(SSI-UM-2.0型,英国Delta公司)测定,选择在晴朗无云天气的11:00—13:00,每个处理选3个点测量LAI,以3次测量值的均值作为该处理的实际LAI,测量频率依天气情况每14 d一次。小麦收获后进行考种,统计有效穗数、籽粒产量和生物量。作物生长速率(Crop growth rate,CGR)通过计算地上部生物量与取样时间(播后时间)线性回归曲线的斜率得到[31]。
1.3.2光合有效辐射
冠层上下的PAR通过安装在冬小麦株间的光合有效辐射传感器(ST-GHY型,陕西盛天伟业电子科技有限公司)进行观测,每个处理各选择一个小区安装传感器,每层高度设置2个不同位置的传感器作为重复。传感器分别安装在地面以上0 cm和120 cm的位置,采样频率为30 min,由配套的数据采集器(ST-CJQ-10型,陕西盛天伟业电子科技有限公司)收集数据,每月下载一次。每日辐射总量通过将辐射传感器的测量值相加后乘以时间步长得到,同时将辐射传感器输出单位光量子通量密度(μmol/(m2·s))转换为能量通量密度(MJ/(m2·s))[32]。冠层的PAR截获量采用HIPPS等[33]提供的方法计算,即
IPAR=Pup-Pbottom
(1)
式中IPAR——冠层的PAR截获通量,MJ/m2
Pup——入射到冠层的PAR辐射通量,MJ/m2
Pbottom——透过冠层到达地面的PAR辐射通量,MJ/m2
PAR的截获率是入射PAR和透射PAR之差与入射PAR的比值,通过RUIZ等[34]的计算方法计算,即
(2)
式中FIPAR——冠层PAR截获率,%
利用FIPAR和LAI,通过比尔定律[35]计算出消光系数
(3)
式中K——消光系数
LAI——叶面积指数
其他气象数据由杨凌国家气象一般站(距离试验田以东约150 m)获得,包括日最高气温(℃)、日最低气温(℃)、日照时长(h)、相对湿度(%)、风速(m/s)。
累积的冠层PAR截获量由冬小麦生育期内每天的截获量相加得到。RUE采用MONTEITH等[5]给出的定义计算,即累积地上生物量与累积冠层PAR截获量的比值。
(4)
式中Ad——地上部生物量,g/m2
D——冬小麦播种后时间,d
全生育期的总RUE是累积地上部生物量和累积冠层截获PAR的线性回归曲线的斜率,线性回归过程中直线不必通过原点[31]。
1.3.3水分胁迫指标
采用土壤相对有效含水率来表征土壤的水分胁迫程度Aw,这是一个无量纲值,取值在0~1之间,0表示完全水分胁迫,1表示无水分胁迫[36-40]。冬小麦根系的主要活动区域是地表以下0~60 cm土层[41],因此本研究选择60 cm深土层的土壤水分数据,以土壤相对有效含水率作为量化各处理冬小麦水分胁迫程度的指标。
(5)
式中θa——土壤体积含水率,cm3/cm3
θwp——凋萎系数,cm3/cm3
θf——田间持水量,cm3/cm3
1.4 统计分析
使用SPSS 23对各处理因素影响进行方差分析;使用LSD法进行差异性检验;使用Excel 2010计算作物生长速率和RUE;使用Matlab 2014a中的曲线拟合工具箱拟合冠层PAR截获率与LAI的关系曲线。
2 结果与分析
2.1 各处理生育期内水分胁迫程度
冬小麦2个生长季的相对有效含水率表现出相同的变化规律。在充分灌水条件下,CK处理的相对有效含水率在整个生育期都高于其余3个处理,受水分胁迫的程度最小,而营养生长期受旱的ES处理其相对有效含水率从播后100 d的100%开始迅速下降,从返青期开始到拔节期结束,其相对有效含水率持续低于CK和LS处理,到拔节期末段趋近于0(图2,G代表进入返青期,J代表进入拔节期,H代表进入抽穗期,F代表进入灌浆期)。而在抽穗期和灌浆期,开始对ES处理进行灌水,但是由于水分胁迫对于作物生长具有滞后效应[42],所以复水后该处理的冬小麦生长仍然受到抑制,其相对有效含水率逐渐增加直至和CK处理持平。生殖生长期受旱的LS处理在返青期和拔节期其相对有效含水率与CK处理有相似变化趋势,平均相对有效含水率在40%以上,从抽穗期开始停止灌水,其相对有效含水率迅速下降,在灌浆期降至最低值并一直持续到收获。WS处理由于生育期内不接受任何水分供应,故其相对有效含水率从出苗到收获逐渐降为0,尤其在返青期后始终处于较低水平。

图2 冬小麦生育期内0~60 cm土层相对有效含水率的动态变化Fig.2 Dynamics of relative available water content for 0~60 cm soil layer during wheat growing season
2.2 叶面积指数和冠层光合有效辐射截获率动态变化

图3 2015—2016和2016—2017生长季不同阶段受旱条件下冬小麦叶面积指数的动态变化Fig.3 Dynamics of winter wheat LAI under water stress at different growth stages during 2015—2016 and 2016—2017 growing seasons
2015—2016年和2016—2017年两季的冬小麦LAI表现出相同的趋势(图3)。在2015—2016年,各处理在返青之前并没有太大的差异。从拔节期开始,CK处理的LAI增速显著高于其他3个处理,特别是在抽穗期,此时营养生长达到顶峰(图3a)。全生育期受旱的WS处理的LAI则始终低于其他处理,这种现象同样出现在2016—2017年的冬小麦试验中(图3b)。在2016—2017年CK、ES和LS处理差异并不明显,但3个处理在抽穗期的LAI大小关系和前一年相同,ES、LS和WS处理最大LAI的2年平均值分别比CK处理低31%、15%和58%,这表明水分胁迫影响了冬小麦的叶片生长。通过比较2年的ES和LS处理可以发现,在冬小麦的返青期和拔节期受旱对其LAI发育影响要比抽穗期和灌浆期大。2年的LAI最大值相差不大,但是2016—2017年冬小麦各处理之间的LAI差异相较于上一年的小很多,这是由于冬小麦生育后期的2017年气温比2016年同期要高,尽管处于受旱条件,但是更多的积温促进了作物的生长[43]。

图4 2015—2016和2016—2017生长季不同受旱阶段冬小麦冠层PAR截获率Fig.4 Fraction of intercepted PAR after greening of winter wheat under water stress at different growth stages of drought during 2015—2016 and 2016—2017 growing seasons
水分胁迫对LAI的影响直接导致了冠层PAR截获率的差异。在低LAI的条件下,冬小麦冠层PAR截获率主要受天气情况和太阳高度角的影响[33],水分胁迫对其影响不大。此外,由于冬小麦从拔节期开始封行,传感器才能准确测量到冠层上下的PAR差异,因此主要分析了拔节期到收获时的冠层PAR截获率的变化情况。在本研究中,2015—2016和2016—2017生长季WS处理的PAR截获率始终低于其他3个处理(图4),而且差异明显。在返青期和拔节期(播后140~180 d),2年的ES处理与CK处理的PAR截获率并无明显差异,而LS处理的PAR截获率均高于CK处理。而在抽穗期和灌浆期(播后180~230 d),ES处理则始终低于CK处理,而LS处理与CK处理无明显差异。CK、ES、LS和WS处理2年的最大冠层截获率平均值分别为90%、88%、79%和42%,这表明冬小麦营养生长阶段受旱对于其PAR截获率有更严重的影响,这与前述研究结论相似。
从以上试验结果可以发现,水分胁迫对冠层PAR截获率的影响主要通过影响叶面积的生长来体现。消光系数描述植物冠层结构以及冠层截获光能力,受种植结构、太阳高度角、叶倾角分布等因素的影响[35]。计算所得CK、ES、LS和WS处理的消光系数分别为0.23、0.29、0.32和0.16(图5),即4个处理的平均值为0.25。而高晓飞等[44]的研究结果显示冬小麦消光系数介于0.50~0.85之间,与之相比,本研究得到的结果要小得多。消光系数在一定程度上反映了大田作物群落结构和受光态势,4个处理中,WS处理的消光系数最小,说明该处理叶片生长很差,冠层透射率高,实际观测中,该处理小麦叶片短小,数量少,水分胁迫对冠层的发育产生了明显的负面影响。消光系数对冠层PAR截获率的估算有直接影响,而冠层PAR截获率能否准确计算对作物干物质的模拟至关重要,但是作物生长模型往往不考虑地理因素、气候条件及管理因素的差别,而将消光系数作为一个常量对待,这可能会使模型模拟结果产生误差。
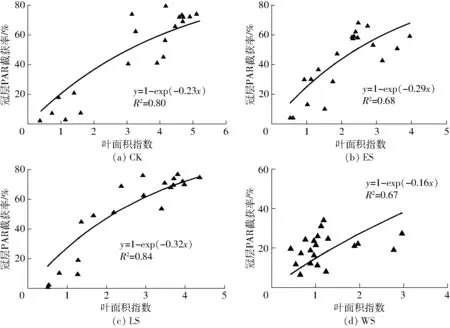
图5 不同受旱阶段处理下冬小麦冠层PAR截获率和LAI的关系Fig.5 Relationship between fraction of intercepted PAR and LAI under different treatments of water stress at different growth stages
2.3 各处理地上部生物量和作物生长速率的比较
本试验中不同处理的冬小麦收获期地上部生物量介于411~1 724 g/m2之间(表2)。2015—2016生长季冬小麦拔节期,WS处理与其他处理的生物量有显著的差异。在抽穗期,ES处理与LS、WS处理相比有显著差异,而与CK没有显著差异。从冬小麦进入抽穗期直到灌浆期开始,营养生长基本停止,CK处理的生物量最大,生物量明显比其他3个处理增加快,同时ES和LS处理的生物量差异不大。从拔节期开始到抽穗期结束,阶段性水分胁迫对于干物质积累影响并不明显。然而,从灌浆期开始一直到冬小麦收获,ES和LS处理的地上部生物量累积速率显著高于WS处理,在收获时,CK处理地上部生物量分别比ES、LS和WS处理高6.5%、6.6%和226.0%。2016—2017生长季的冬小麦生物量和上一生长季有相似的趋势,冬小麦收获期地上部生物量CK处理比ES、LS、WS处理高10.3%、11.3%、160.4%。在收获期,CK、ES、LS和WS处理2年的平均地上部生物量分别为1 532、1 410、1 403、537 g/m2。本研究表明全生育期受旱显著影响冬小麦地上部生物量的积累,而在冬小麦的某个发育阶段受旱,对其生物量累积并不会有显著差别。2016—2017生长季的最终地上部生物量要显著高于2015—2016生长季,主要原因可能是因为2016—2017生长季的冬小麦生育期内日平均气温较高,冬小麦生育期内有更高的积温[45-46]。
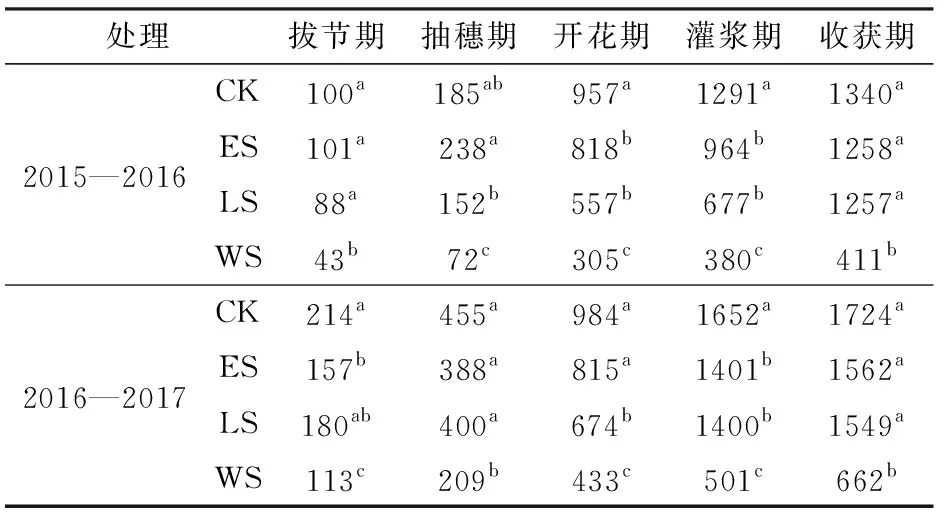
表2 2015—2016和2016—2017生长季不同处理下冬小麦各生育期地上部生物量Tab.2 Aboveground biomass of winter wheat under different treatments of water stress at growth stages in 2015—2016 and 2016—2017 growing seasons g/m2
注:每一列的不同字母代表同一年的不同处理间的差异显著(P<0.05)。下同。
研究中4种处理的作物生长速率与地上部生物量表现出相同的规律(表3)。地上部生物量和作物生长速率之间相关性较好(图6)。充分灌水的CK处理的作物生长速率显著高于其他3个处理,最低的是全部受旱的WS处理,而ES和LS处理之间差异不大,CK处理的作物生长速率分别比ES、LS以及WS高13%、15%和222%。整个试验中,全生育期受旱和生殖阶段受旱对冬小麦的生长速率负面作用更大。

表3 2015—2016和2016—2017生长季不同处理下冬小麦的作物生长速率Tab.3 Crop growth rate of winter wheat under different treatments during 2015—2016 and 2016—2017 growing seasons g/(m2·d)

图6 地上部生物量与作物生长速率的关系Fig.6 Relationship between aboveground biomass and crop growth rate
2.4 不同处理的RUE变化趋势
辐射利用效率RUE被定义为地上部累积生物量与累积PAR截获量的比值[47-48]。在2个生长季中,营养生长阶段受旱的ES处理始终低于生殖生长阶段受旱的LS处理,同时也低于对照处理(图7)。LS处理在拔节期的RUE高于对照处理,进入开花期后,LS处理的RUE开始低于对照处理,这种关系一直持续至小麦收获(图7)。2016—2017生长季的冬小麦RUE整体要比2015—2016生长季高一些(图7),这主要是因为2016—2017冬小麦生长季期间更高的日平均气温引起的,与ANDRADE等[47]的研究结果相似。冬小麦的RUE在整个生育期呈现先增大后减小的趋势,这与之前研究将整个生育期RUE视为常数并不一致。除此之外,RUE在每年的开花期达到最大,4个不同处理(CK、ES、LS、WS)的2年平均值分别是4.77、3.34、3.90、3.87 g/MJ。相比CK处理,水分胁迫确实减低了冬小麦的RUE。在2个生长季WS处理的RUE呈现出不同的变化规律(图7),在2015—2016生长季,WS处理低于其他处理,而在2016—2017生长季,WS处理则仅低于对照处理,这说明与阶段性受旱相比,冬小麦在全生育期持续受旱的情境下有不同的响应机制。

图7 2015—2016和2016—2017生长季不同受旱处理下冬小麦的RUE的动态变化Fig.7 Dynamics of RUE under different treatments of water stress during 2015—2016 and 2016—2017 growing seasons
通过对累积地上部生物量与累计PAR截获量的线性回归,得到每年的总体RUE。在2年的冬小麦受旱试验中,冬小麦的累积地上部生物量与冬小麦冠层的累积PAR截获量有很好的相关性(图8)。WS处理的冠层的累积PAR截获量在2年的试验中平均比CK、ES、LS减少61%、61%和56%。整个生育期的RUE仍然是CK处理最大,为3.55 g/MJ(图8),ES、LS和WS处理的RUE分别比CK处理低22%、5%和22%。此外2年营养生长期受旱的ES处理总体RUE平均值比生殖生长期受旱LS处理减少18%,说明营养生长阶段施加水分胁迫对于干物质的积累比在生殖生长阶段更加不利。2年的WS处理的RUE与ES处理接近,这主要是由于2年的WS处理RUE较大的差异相互抵消所致。
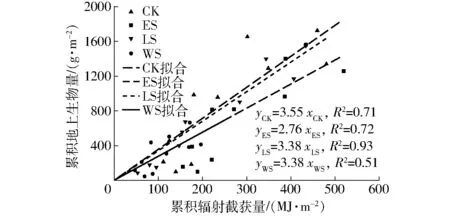
图8 冬小麦累计生物量和累积辐射截获量的关系Fig.8 Relationship between cumulative aboveground biomass and cumulative intercepted PAR of winter wheat
2.5 产量分析
两个生长季内冬小麦各处理之间千粒质量均没有显著的差异,但是CK处理的单株穗粒数显著高于其他3个处理。2015—2016年CK处理的单位有效穗数比ES、LS、WS高21%、27%、31%,而2016—2017生长季的单位有效穗数却没有大的差异(表4)。在籽粒产量方面,2015—2016生长季冬小麦的CK和ES处理显著高于其余2个处理,2016—2017生长季各处理间具有相似的规律,但是CK、ES、LS处理间差异未达到显著性水平, 在2个生长季内WS处理的产量要远低于其他处理(表4)。生殖生长期受旱的处理其籽粒产量和有效穗数均低于营养生长阶段受旱的处理,表明灌浆阶段的水分胁迫对冬小麦产量的形成更加不利。
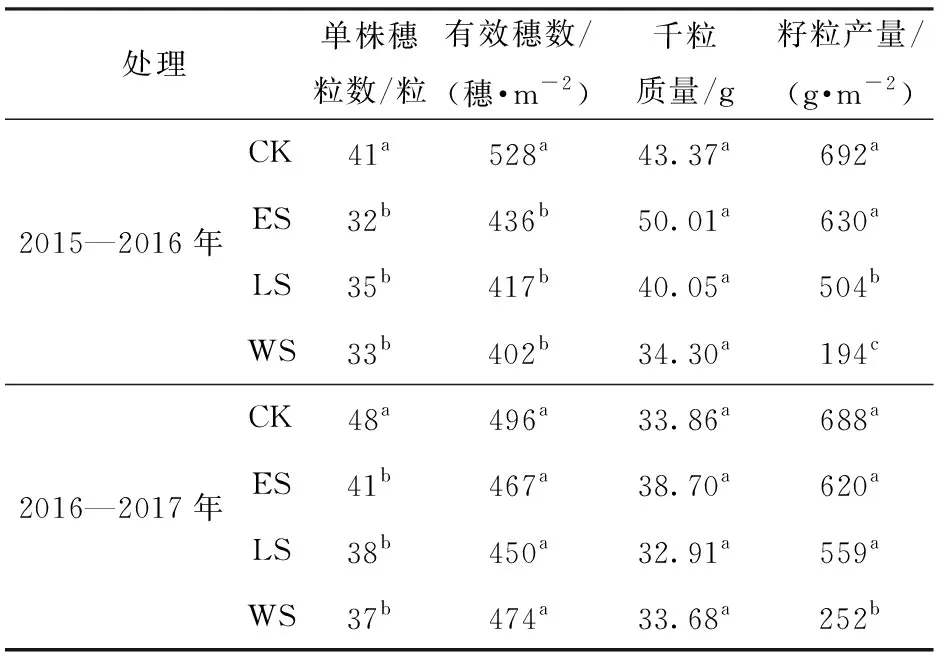
表4 2015—2016和2016—2017生长季冬小麦籽粒产量和产量构成Tab.4 Grain yield and yield components of winter wheat in 2015—2016 and 2016—2017 growing seasons
3 讨论
本研究中,冬小麦遭受水分胁迫的时段是影响冬小麦LAI、冠层PAR截获率和RUE变异性的主要因素。不同处理地上部生物量的差异主要由冠层PAR截获率的差异引起,而冠层PAR截获率的差异则主要由不同受旱阶段产生的LAI的差异导致[18,31,49]。
整个试验过程中,CK、ES、LS和WS 4个处理的最大冠层PAR截获率分别为90%、88%、79%和42%。类似的稀疏小麦冠层,低截获率(小于50%),低叶面积指数(小于1.5)试验现象也出现在半干旱气候的澳大利亚西南部[50-51]。与此相对,BORSTLAP等[52]观测到雨养小麦的截获率高达95%。本试验开始前已经进行了2年相同的受旱试验,LS处理所在的小区土壤墒情比CK处理所在小区差(图2),导致LS处理的LAI略微低于CK处理,但是相比于CK处理,遭受轻微水分胁迫的LS处理冠层叶片分布更加均匀,结构更加有利于截获辐射,因此在该阶段LS处理的冠层PAR截获率要高于CK处理。本研究最大的冠层截获率出现在CK处理中,试验数据表明冠层PAR截获率比生物量、作物需水量等能更好地反映水分胁迫对作物生长的影响。
冠层截获率和LAI随时间的变化规律常被用来求解消光系数[53]。本研究中综合2年的LAI和冠层PAR截获率求出的消光系数分别为0.23、0.29、0.32和0.16(图5)。不同处理间的消光系数差异显著,与高晓飞等[44]的结果相似。WS相较于其他处理,消光系数要小得多,表明冠层稀疏,叶片短小,这符合实际观测的结果。LS处理消光系数高于ES处理,表明生殖生长期受旱对冬小麦叶片生长影响低于营养生长期,这与LAI的结果类似。然而,高晓飞等[44]的研究结果表明冬小麦消光系数介于0.50~0.85之间,本研究结果总体偏小,主要是由于小麦品种、种植密度、种植方式以及测量时间的差异引起的[12,44,54]。在不同的作物模型中,消光系数取值有很大的差异。CERES-Wheat模型中小麦的消光系数是0.85[55],而EPIC 和ALMANAC等许多模型中小麦消光系数是0.65[56-57]。而本研究结合前人的许多研究发现,受不同的作物基因型[12]、种植密度[44]、群体分布[54]影响,消光系数是在一定范围内不断变化的,甚至在作物生长的不同生育期内,消光系数也是不同的[44]。消光系数取值的差异直接会影响冠层PAR截获率的计算,进而影响作物模型对光合作用同化速率的估算。因此,如果能够进一步理解水分胁迫对消光系数的影响机制,将有助于提高作物模型模拟干旱胁迫条件下作物产量的模拟精度,而本研究的结果能够为作物模型在这方面的改进工作提供一定的理论参考。
累积的冠层PAR截获量是绿色冠层持续时间和冠层PAR截获率共同作用的结果[58]。本研究中冬小麦的最终RUE介于2.76~3.55 g/MJ之间,这个结果高于O’CONNELL等[20]报道的1.81 g/MJ,也比VERSTEEG等[59]记录的RUE取值范围(1.7~1.8 g/MJ)要高。本研究采用地上部生物量计算RUE时并没有去除已经衰老的叶片,尽管这部分生物量很小,但依旧会高估总的地上部生物量。在2个生长季WS处理的RUE表现不稳定,尤其在2016—2017生长季该处理RUE很高,产生这种现象的可能原因主要有:①WS处理由于全生育期遭受严重的水分胁迫,整个生育期LAI都保持很低的值,这导致了冠层无法完全闭合,在低冠层郁合度条件下,冠层PAR截获率受天气状况和太阳高度角影响很大,传感器无法有效获取冠层上下的PAR值,很容易低估冠层的PAR截获量,因此针对该处理的PAR截获率的测量需要改进试验方案。②由于WS处理生育期内始终处于严重水分胁迫状态,其生育进程会加快,相比其他处理,更早的进入生殖生长阶段,更多的植物组织提前衰老(衰老的组织不参与干物质的分配),但是在测定地上部生物量时并没有去除这部分衰老的组织,导致高估了地上部生物量。
各处理之间,ES、LS和WS处理的RUE均低于CK处理,水分胁迫对RUE的影响显著,充分灌水的冬小麦有更高、更稳定的光转化效率。RUE的差异反映了水分亏缺、养分胁迫、疾病以及低温等环境因素对冬小麦生长发育的影响。产生这种差异的原因也有2个:①不同阶段水分胁迫下作物根部和地上冠层光合同化物分配的差异造成。②由不同阶段水分胁迫下PAR截获的差异引起[20]。相同的气候条件下,冬小麦为了适应水分胁迫,会将更多的光合同化物分配至根部,导致根的活动区域超出60 cm,这会产生更大的根冠比[60-61],进而导致更低的RUE。根据KINIRY等[62]的研究,冬小麦的根生物量在开花期能超过总的生物量的10%,而INCERTI等[63]观测到的这个数据甚至高达46%。因此,进一步研究水分胁迫条件下作物根冠比的动态变化对于精确计算RUE十分重要。
已有研究认为作物整个生育期的RUE是一个常量[48],而在本研究中发现冬小麦的RUE从出苗到收获呈现先增大后减小的趋势,RUE在每年的开花期达到最大,返青期和灌浆期变小。类似的变化规律BRODRICK等[64]在高密度种植的棉花实验中发现过,苜蓿在生长过程中RUE的变化也有一样的趋势[65],生育期的开始阶段和结束阶段RUE较低,生长中期RUE高。这种现象也符合冬小麦在拔节到开花期间生长不断加快的常识。从拔节期开始到开花期,作物生长速率加快,尤其是水分充足条件下,短期内更高的RUE能够产生更高的生物量。
作物生长模型作为研究作物生理过程的一种有效工具,而逐日同化产物的形成模拟是其中重要的一环[7, 66-67]。作物模型中群体的日同化量大都表示为冠层PAR截获率与RUE的函数[44]。入射PAR是位置、时间、播期和作物物候期的函数,而冠层PAR截获率是LAI和消光系数的函数。作物生长模型能否准确模拟作物干物质形成取决于其对LAI、消光系数以及RUE的充分描述。作物生长模型通常不考虑地理因素、气候条件以及管理因素而直接为每种作物分配一个RUE,如CERES-Wheat模型的小麦的RUE默认是2.7 g/MJ,CERES-Maize模型中玉米的RUE默认为4.1 g/MJ[6],小麦的RUE略微低于本研究的结果,而且模型中RUE也未考虑水分胁迫、养分胁迫、种植密度和群体结构等因素的影响。此外,由于RUE在作物的整个生育期是一个动态变化的参数,在作物模型计算不同生育期的干物质量时使用恒定的RUE会产生误差,这对于作物模型的准确性是不利的。综上,为了作物模型更加精确地模拟水分胁迫条件下的作物生长和产量形成过程,建议应将消光系数和RUE作为生育期或者积温的函数来对待而非单一常数,需进一步考虑水分和氮素胁迫对消光系数和RUE的次级影响。
4 结论
(1)当冬小麦不同生育期发生水分胁迫时,其冠层结构和截获PAR的能力均受到一定影响,LAI显著降低,消光系数也比正常灌水处理小得多。水分胁迫对冬小麦冠层PAR截获率产生明显的负面影响。此外,营养期受旱对冬小麦冠层PAR截获率减少的影响大于生殖期受旱。
(2)冬小麦RUE介于2.76~3.55 g/MJ之间,比已有研究略高,但阶段性水分胁迫降低了RUE,尤其是在营养期受到水分胁迫的影响更大。
(3)冬小麦RUE在整个生育期不是一个常量,而是保持先增大后减小的动态变化趋势,在开花期时冬小麦的RUE达到最大。要想作物模型更加精确地模拟水分胁迫条件下的作物生长和产量形成过程,建议将消光系数和RUE作为生育期或者积温的函数来对待而非单一常数,并应进一步考虑水分和氮素胁迫对消光系数和RUE的次级影响。
