黑土区水稻生长生理特性与产量对耗水过程的响应
2018-09-17魏永霞杨军明侯景翔
魏永霞 汝 晨 吴 昱 刘 慧 杨军明 侯景翔
(1.东北农业大学水利与土木工程学院, 哈尔滨 150030; 2.东北农业大学农业部农业水资源高效利用重点实验室, 哈尔滨 150030; 3.黑龙江农垦勘测设计研究院, 哈尔滨 150090; 4.东北林业大学林学院, 哈尔滨 150040; 5.东北农业大学理学院, 哈尔滨 150030)
0 引言
我国是世界上最大的水稻生产国,水稻种植面积占粮食总面积的28%,耗水量占全国总用水量的54%左右,占农业总用水量的70%左右[1]。东北地区水资源在年际间、地区间以及年内分布极不均衡,严重制约了东北地区水稻的生产发展。因此,发展高效的节水灌溉技术对于东北黑土区农业生产应对水资源短缺及农业可持续发展具有重要意义。
节水的有效途径就是根据水稻不同时期对水分敏感性进行针对性的水分调控,从而使得水资源得到有效利用[2]。水稻耗水过程是指在全生育期内水稻维持自身生长发育、生理活动所需水量的动态变化[3],水稻生长生理对水分的响应和适应规律一直是生物逆境学研究的重点。近年来有关研究表明,适度干旱胁迫有助于构建适宜株型结构,提高叶片活力和净光合速率,为水分高效利用和水稻优质高产奠定了基础。邵玺文等[4]认为,在反应敏感阶段保持浅水层能够增强后期根系活力,延缓叶片衰老时间,增加干物质积累量。大量研究表明,水分胁迫对水稻干物质积累[5-6]、根系活力[7]、光合生理特性[8-9]也存在显著影响,良好的土壤水分条件是水稻正常生长与高产的基础。段素梅等[6]研究则认为,在某一阶段适度亏水可使复水后短期作物生长生理特性得以改善,不同生育期水稻受水分胁迫的影响机理不同,这体现了水稻对环境水分变化具有一定的适应性。以往研究中,多关于水分胁迫对水稻产量形成的影响,主要集中在单一阶段、同等水分胁迫程度下的研究,缺乏科学系统的比较分析,针对精准调控条件下水稻生长生理特性对耗水过程响应的研究较少。因此,本试验开展水稻耗水过程对水稻生长、生理特性及产量影响的研究,分析水稻生长生理特性及产量对水分条件变化的响应特征与规律,旨在探索不同阶段控水带来的正负效应及缓解这种影响的措施,有助于深入了解水稻对水分逆境的抗性机理,充分挖掘水稻生产潜能,以期为黑土区水稻高产、节水栽培提供依据。
1 材料与方法
1.1 研究区概况
试验区位于黑龙江省庆安县和平灌区水稻灌溉试验站基地(北纬46°41′~47°4′,东经127°20′~127°49′),属于北温带半干旱、半湿润的大陆性气候。多年平均降水量550 mm,多年平均温度2.5℃,全年无霜期128 d。试区土壤种类为白浆型水稻土,土壤基础理化指标:容重为1.02 g/cm3、孔隙度61.6%、0~30 cm体积饱和含水率平均为55.5%、pH值为6.51、有机质质量比41.5 g/kg、全氮质量比15.10 g/kg、全磷质量比15.21 g/kg、全钾质量比20.09 g/kg、碱解氮质量比153.98 mg/kg、有效磷质量比25.21 mg/kg、速效钾质量比157.19 mg/kg。
1.2 试验设计
试验在HSY.XHZ-1型自动称重式蒸渗仪(测筒高1.5 m,直径1.13 m,表面积1 m2,测量精度100 g)内进行,供试水稻品种为龙庆稻3号,插秧规格为行株距22.5 cm×12.5 cm,共4行24穴,每穴定5株,并配备移动式遮雨棚,防止降雨的干扰。蒸渗仪四周种植同一品种水稻作为保护田。根据作物各生育阶段长度接近等原则,以水稻6个生育时期耗水量为试验因素,采用U7(76)均匀试验设计,在试验范围内挑选出具有代表性的试验点,这些点在试验范围内充分均衡分散,并能够反映体系的主要特征。60%土壤饱和含水率是水稻生长发育相对较低的水分控制标准,设置60%~100%的水分控制标准为重度调亏,依次较均匀地选取重度调亏与适宜灌水下限间的不同田面水层深度和土壤湿度,直至设置20~30 mm的田面水层深度为水分控制的最高标准,即完成6个土壤水分控制标准的选取,而对照处理(CK)是参照当地大田的水分控制标准进行设计,同时也保证了均匀设计中各时期不同处理间控水标准存在一定差异,从而可探究各因素对指标的影响及指标的变化规律。共7个处理,3次重复,21个蒸渗仪。各处理水稻各生育时期水分调控标准见表1。各处理均采用相同田间管理方法,施肥、除草等均保持一致,全生育期总施氮110 kg/hm2,基肥∶分蘖肥∶促花肥∶保花肥比例为4.5∶2∶1.5∶2;施P2O545 kg/hm2,全部用于基肥一次性施入;施K2O 80 kg/hm2,分基肥和水稻8.5叶龄时期(幼穗分化期)两次施入,比例为1∶1。于2017年5月18日插秧,9月20日收获。

表1 水稻不同生育期水分控制标准Tab.1 Water control standards at different growth stages of rice
注:“%”为占土壤饱和含水率的百分比,“mm”为水层深度。
1.3 观测指标与方法
1.3.1作物耗水量
每天08:00采用便携式土壤水分速测仪观测蒸渗仪内土表的水分变化(无水层),采用专用水尺测定水层深度的变化(存在水层);土壤水分到达控制下限即灌至设计上限,灌水前后需加测,灌水量由水表读出。并于每天08:00使用称量式蒸渗仪获取总体质量,根据前后的质量差(W1)计算水稻每天实际蒸散量(耗水量),由于试验在自动感应式遮雨棚内进行,且蒸渗仪底部有底,故不考虑生长季内降雨量、地表径流量、地下水补给量的影响。当在各个生育时期对植株干物质积累量进行取样时,需记录取样前、后蒸渗仪的总体质量,两次质量差值(W2)为取样引起蒸渗仪的质量变化,则W1-W2的差值即为各生育时期取样当天的耗水量。作物阶段耗水量、耗水强度、耗水模系数计算公式为
ETi=ΔWi+Ii
(1)
CRi=ETi/di
(2)
Ri=ETi/ET×100%
(3)
式中ETi——阶段耗水量,mm
ΔWi——阶段储水量差值,mm
Ii——阶段灌水量,mm
CRi——阶段耗水强度,mm/d
di——阶段持续时间,d
Ri——阶段耗水模系数,%
ET——全生育期耗水总量,mm
i——生育阶段,i为1、2、3、4、5、6分别代表分蘖前期、中期、后期,拔节孕穗期、抽穗开花期和乳熟期
1.3.2水稻各器官干物质积累量
分别于分蘖前期至成熟期,每处理选取代表性植株3穴,将样品地上各部分(叶片、茎鞘、穗)擦拭表面灰尘后分别装袋,同时进行根系取样,挖取长等于行距、宽等于株距、深40 cm带根土块,用流水缓慢清洗,直至泥土冲洗干净为止,洗净后用无氮吸水纸吸干装入袋中,把各时期样品装袋后于105℃杀青30 min,调至80℃干燥至恒质量,冷却后用精度0.01 g电子秤测定。
1.3.3根系伤流量
分别于分蘖前期至乳熟期每个处理选取代表性植株3穴,剪去土面10 cm以上的地上部植株,套上已称量装有脱脂棉的自封袋密封(每蘖1袋),并用橡皮筋将其扎紧。当天18:00开始套袋,第2天8:00将其取下称量,收集后与收集前质量差值即为单茎伤流量。
1.3.4叶片光合指标
每个蒸渗仪选取3个测试样点进行观测,在各生育时期对已标记的代表性单株功能叶进行测定,采用美国LI-COR公司生产的LI-6400型光合仪在每个生育期的10:00—11:00测定功能叶片净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci),数据由仪器自动读出。
1.3.5测产
成熟期每个蒸渗仪选取6穴代表性水稻单打单收计算每穴实际产量,根据每公顷穴数计算每公顷产量。
1.4 数据处理方法
以分蘖前期、分蘖中期、分蘖后期、拔节孕穗期、抽穗开花期、乳熟期的耗水量为变量,采用多元回归分析与各因变量建立水分响应模型,并对模型及各因素进行显著性检验;运用Excel进行数据处理,Origin 9.10进行作图,SPSS 22.0作回归统计分析;多重比较采用LSD法,显著水平取0.05。
2 结果与分析
2.1 水稻各生育时期耗水特征
表2为不同水分处理下水稻各生育时期的耗水特征。从整个生育期来看,水稻各处理生育期内耗水强度总体呈现先稳定上升后逐步下降的趋势,6月中旬以前,日平均气温较低致使分蘖前期历时稍长,植株蒸腾量较小,但水稻耗水强度仍然较高,在2.87~3.74 mm/d之间;分蘖中期水稻进入旺盛的营养生长阶段,田间耗水由棵间蒸发为主转向植株蒸腾为主,至分蘖后期平均耗水强度逐渐增加至4.56 mm/d。随着生育进程推进和高温天气的持续,抽穗开花期平均耗水强度增至5.66 mm/d,此时期是水稻营养生长和生殖生长并进阶段,植株生长旺盛且冠层覆盖率较高,水稻需水量大。水稻灌浆后,各处理耗水强度开始迅速回落,至乳熟期耗水强度降低为3.56 mm/d。各处理耗水模系数在整个分蘖期最大,平均值为43.98%,抽穗期后耗水模系数基本趋于稳定,乳熟期各处理耗水模系数均值为16.02%,此时期是籽粒灌浆、产量形成的重要阶段,故耗水模系数同样较高。
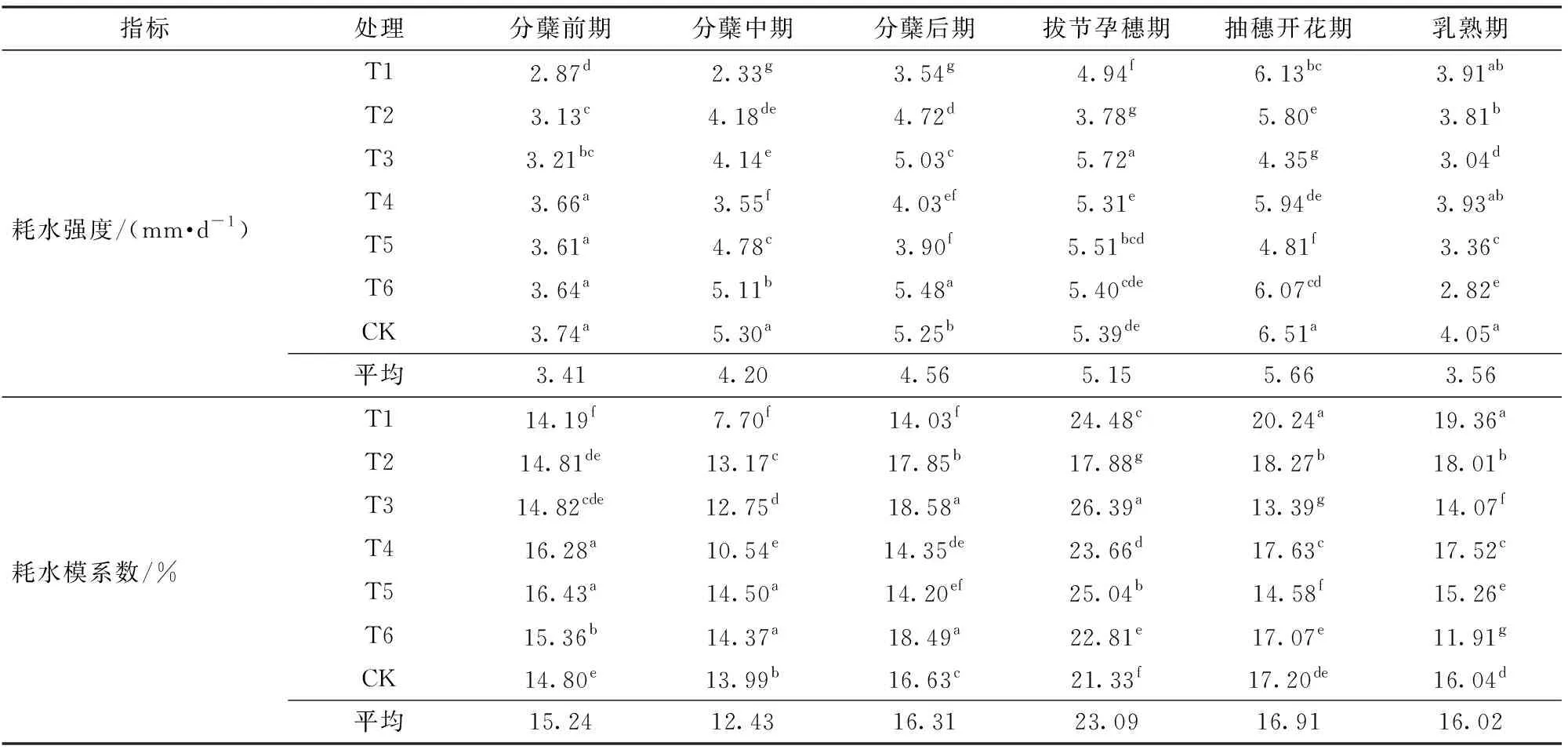
表2 各生育期耗水特征Tab.2 Water consumption characteristics of each growth stage
注:同列不同小写字母表示在0.05水平上差异显著。
2.2 耗水过程对水稻各器官干物质积累的影响
图1为不同处理各生育时期水稻各器官干物质积累的变化曲线。从图1可知,不同处理水稻各器官干物质呈现出相同器官具有相同的变化规律。水稻叶片干物质量随着生育进程的延续均呈现单峰曲线变化趋势,叶片干物质量在拔节孕穗期达到最大。茎鞘干物质量在抽穗开花期达到最大,此时期茎鞘干物质最大积累量可为后期光合产物向籽粒转移提供有力保障。而水稻叶片、茎鞘干物质积累量在达到最大值后开始集中流向穗部,使穗干物质量在抽穗开花期后迅速增加。各处理根部干物质量的变化与叶片、茎鞘呈现相似的规律,且呈倒“V”型曲线,其在抽穗开花期达到最大。

图1 不同生育期水稻各器官干物质积累动态曲线Fig.1 Dynamic curves of dry matter accumulation of rice various organs at different growth stages
由于各处理在不同生育阶段控水标准不同,导致水稻各器官干物质积累过程呈现出一定差异。分蘖中期开始叶片干物质量差距逐渐显现,至分蘖后期各处理叶片干物质量极差达到最大,为3.4 g/穴;T3、T5处理在分蘖后期至拔节孕穗期叶片干物质量增长幅度较CK处理分别提高40.3%、51.8%,可能是由于旱后复水给拔节孕穗期叶片生长带来一定的超补偿作用;T1、CK处理在拔节孕穗期后叶片干物质量降低趋势相对平缓。分蘖中期各处理茎鞘干物质量增加幅度为全生育期最大,为5.08~7.7 g/穴。分蘖后期T2、T4处理的茎鞘干物质量增加幅度分别为5.45、6.02 g/穴,均高于CK处理;抽穗开花期进行水分调控的各处理茎鞘干物质量增加幅度均低于CK处理。拔节孕穗期穗干物质量差异不明显,T1处理在抽穗开花期进行复水,但其穗干物质量仍然最低,为6.44 g/穴。说明抽穗开花期前的连续亏水使水稻营养器官发育受阻,进而对生殖生长阶段的籽粒灌浆过程带来不利影响。分蘖前期、分蘖中期以CK处理的根干物质量最大,分别为1.67、4.24 g/穴,分蘖后期开始,进行水分调控的个别处理根干物质量高于CK处理,T3处理的抽穗开花期根干物质量增加幅度最低,为2.17 g/穴。
水稻各器官干物质积累量是反映作物生产能力的一个重要指标,且产量的高低取决于光合物质积累量及其分配和转化是否协调合理。水稻籽粒的灌浆物质大部分来自于抽穗期前后叶片、茎鞘、根部的贮积和抽穗后转运到穗部的非结构性碳水化合物[10]。因此,探究不同时期耗水量对各器官干物质最大积累量的影响,对于提高各器官干物质量以将更多营养物质转向穗部籽粒从而提高产量具有重要意义。分别建立叶片最大干物质量(ya)、茎鞘(yb)及根最大干物质量(yd)、穗最大干物质量(yc)与耗水量(分蘖前期至对应时期)的水分响应方程,如表3所示。各方程均达到显著或极显著水平,决定系数R2均较高,表明各方程拟合效果良好。

表3 水稻各器官干物质积累水分响应方程Tab.3 Moisture response equation of dry matter accumulation in various organs of rice
通过方程ya可知,ET1未通过显著性检验(P1=0.089),其余变量达显著或极显著水平(P2=0.017,P3=0.002,P4=0.031)。分蘖前期作物需水量低,ET1不会对ya造成显著影响。其中ET3系数最大,说明此时期供水对ya的影响最大。由方程yb可知,ET1、ET3未通过显著性检验(P1=0.063,P3=0.067),其余变量达显著或极显著水平(P2=0.038,P4=0.025,P5=0.004)。其中ET2系数与ET4相差不大,说明两个生育时期耗水量对水稻茎鞘干物质量影响程度较为接近;ET5系数最大,说明抽穗期耗水量对yb影响最大。从方程yc可知,各变量均通过显著性检验,且均对yc呈显著或极显著正影响(P1=0.035,P2=0.004,P3=0.037,P4<0.001,P5<0.001,P6=0.029)。ET2系数为0.095,在0.01下达极显著水平,表明分蘖中期耗水量对yc存在极显著影响。由方程yd可知,ET1、ET5未通过显著性检验(P1=0.081,P5=0.056),其余变量均达显著水平(P2=0.034,P3=0.016,P4=0.024)。ET2系数为0.04,表明分蘖中期耗水量对yd存在显著正影响。ET3、ET4的系数为负,表明分蘖后期、拔节孕穗期耗水量均对yd存在显著负影响。
2.3 耗水过程对水稻根系伤流量的影响
图2为不同处理各生育时期水稻根系伤流量的动态变化曲线。从图2可以看出,水稻各处理根系伤流量在生育期内呈现单峰曲线的变化趋势,根系伤流量在抽穗开花期达到最大值,乳熟期下降幅度明显。建立抽穗开花期根系伤流量(y)与该指标达到最大值前各生育阶段耗水量的水分响应方程
y=1.401+0.001ET1+0.005ET2-0.004ET3-
0.003ET4-0.003ET5(R2=0.921,P<0.01)
(4)

图3 抽穗开花期各处理叶片气孔导度、胞间CO2浓度、蒸腾速率及净光合速率Fig.3 Leaf Gs, Ci, Tr and Pn of different treatments at heading-flowering stage
方程在0.01条件下达极显著水平,决定系数R2较高,表明该方程能够良好地反映根系伤流量与阶段耗水量之间的关系。由方程y可知,ET1未通过显著性检验(P1=0.067),ET2、ET3达极显著水平(P2=0.005,P3=0.007),ET3、ET4达显著水平(P4=0.018,P5=0.019),其中ET3、ET4、ET5对y为负效应。ET2系数最大,表明分蘖中期耗水量对y的调控效应最大;而ET3、ET4、ET5均对y起到一定负作用,这是因为分蘖中期根部器官已获得良好发育,分蘖后期至抽穗期的水分胁迫虽然会导致土壤含水率低,水稻生长对恶劣环境产生的“抗性”和根系生长的趋水性促进受旱时稻根向深处延伸,特别是促进了吸收根的生长,增强根系的吸收能力[11]。抽穗开花期后水稻各处理根伤流量呈下降趋势,这可能由于根系衰老和部分根系死亡的缘故。

图2 不同生育期水稻根系伤流量动态曲线Fig.2 Dynamic curves of bleeding amount of root at different growth stages
2.4 水稻叶片光合特性对耗水过程的响应及其机制分析
2.4.1耗水过程对水稻叶片光合特性指标的影响
光合作用是光合物质生产的源泉,气孔导度、净光合速率、蒸腾速率和胞间CO2浓度等都是表征群体光合生产能力的重要指标。不同处理水稻叶片的气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr)及净光合速率(Pn)均在抽穗开花期达到最大,探究耗水过程对抽穗开花期叶片光合特性的影响,对于增强抽穗期后光合生产物质的积累及更多地向穗部转运提供重要依据[12]。图3为抽穗开花期叶片的各光合特性指标,从图3可知,T3处理Gs最低,为0.30 mol/(m2·s),与T1、T2处理均未达到显著性差异(P>0.05)。T2处理Ci最低,为325.32 μmol/mol;T4、T5处理间Ci差异不显著,但均显著高于其他亏水处理。T6处理Tr高于T4处理,差异不显著,但显著低于CK处理(P<0.05);各处理与T3处理之间Tr均达到显著性差异(P<0.05)。T1、T2、T3处理之间Pn差异不显著,且均处于较低水平;而T4处理Pn均显著高于其他亏水处理(P<0.05)。整体来看,CK处理的4种光合指标均显著高于其他处理(P<0.05)。分别将抽穗开花期Pn、Gs、Ci及Tr与相应各时期耗水量建立水分响应方程,如表4所示。对各方程进行显著性检验,均在P<0.01条件达极显著水平,其决定系数R2均在0.80以上,表明各方程的拟合效果良好。

表4 各光合特性指标水分响应方程Tab.4 Moisture response equation of different photosynthetic characteristics indexes
气孔导度反映了气孔对外界气体及水蒸气的传导程度,是与光合作用密切相关的一个指标。由方程Gs可知,ET1、ET3未通过显著性检验(P1=0.075,P3=0.052),其余变量均达到显著或极显著水平(P2=0.037,P4=0.016,P5<0.001)。ET2相比ET1、ET3系数大,表明分蘖中期耗水量比分蘖前期、分蘖后期对Gs影响更为显著;ET4、ET5对Gs均存在显著正影响,但Gs对ET5的响应最为敏感。
胞间二氧化碳浓度的变化反映了叶肉细胞光合作用能力,可以作为判断影响光合速率变化主要原因的依据。由方程Ci可知,ET1未通过显著性检验(P1=0.089),其余变量均达到显著或极显著水平(P2<0.001,P3=0.021,P4=0.009,P5<0.001),其中ET3对Ci存在显著负效应。ET2系数高于ET4,表明分蘖中期耗水量比拔节孕穗期对Ci影响更大;ET3系数为负值,表明分蘖后期耗水量对Ci有显著负影响。ET5系数最大,这说明植株从分蘖前期保持良好的生长发育至抽穗开花期时,ET5才是对Ci影响最显著的变量。
蒸腾速率不仅受外界环境条件的影响,而且还受植物本身的控制和调节,是一种复杂的生理过程。由方程Tr可知,ET3未通过显著性检验(P3=0.072),其余变量均达到显著或极显著水平(P1=0.028,P2=0.019,P4<0.001,P5<0.001),ET1对Tr有显著负效应。随着生育时期推进,作物耗水量逐渐成为制约叶片蒸腾速率的主要因素,ET2、ET4、ET5对Tr均达到显著正效应,其中ET4、ET5系数较高分别为0.061、0.073,表明拔节孕穗期、抽穗开花期耗水量对Tr的影响较其他时期更为显著。
净光合速率是表征植物光合作用能力的一个重要指标,同时也是物质积累和产量形成的基础[13]。由方程Pn可知,ET1未通过显著性检验(P1=0.092),其余变量对Pn的影响均达到显著或极显著水平(P2=0.001,P3=0.038,P4=0.009,P5=0.004)。分蘖前期水稻营养器官尚未发育成熟,叶片光合能力较弱,分蘖前期耗水量对Pn的影响不显著。ET2系数较ET4、ET5较大,表明分蘖中期耗水量对Pn的正效应最大,也说明了分蘖中期供水充足条件下叶片良好生长对Pn的提升具有重要保证。而ET3系数为负,表明分蘖后期耗水量对Pn有显著负效应。
2.4.2净光合速率对气孔导度、胞间CO2浓度及蒸腾速率的响应关系及其机理分析
抽穗开花期各处理的Gs、Ci、Tr、Pn之间的拟合效果如图4所示,各拟合方程的R2均较高,表明方程能够较好反映各光合特性指标之间的关系。由图4可知,叶片气孔导度能直接或间接对胞间CO2浓度及蒸腾速率产生影响,叶片Ci随着Gs的增加呈二次函数关系,而当叶片Gs降低时Ci也随之降低,而后趋于平缓。Tr与Gs呈二次函数抛物线关系,Tr随着Gs增加而增加,但其增加幅度不断降低;由图4c、4d可以发现,胞间CO2浓度、蒸腾速率与净光合速率也存在函数关系,Tr与Pn呈线性正相关关系,叶片Pn随着Tr的增加呈直线增加趋势;而叶片Pn随着Ci增加呈抛物线趋势。

图4 抽穗开花期光合特性指标间拟合效果Fig.4 Fitting charts of photosynthetic characteristics indexes at heading-flowering stage
分析Gs、Ci、Tr与水稻耗水过程响应关系可以进一步明确Pn对耗水过程的影响机理,结合表4和图4分析可知,分蘖前期植株生理活动较弱,ET1对Gs影响未达显著水平(P>0.05),不会引起外界CO2供应受阻,进而对Pn无显著影响。ET2对Pn调控效应最大,此时期耗水量会显著影响抽穗开花期Gs,阻碍了CO2的供应进而导致Ci减小,会引起抽穗开花期Pn降低。ET3对Pn存在一定负影响,这是由于水稻对环境CO2扩散至细胞阻力很小,这种低气体扩散抗阻特性使得叶片在部分气孔关闭同时,依然能保持较高胞间CO2浓度供给,保证光合作用进行[14],另外分蘖后期亏水会对生育后期复水产生一定超补偿作用,一定程度引起抽穗开花期Ci增大,对Pn增加奠定良好基础[15]。ET4增加会显著提升Pn,结合图4a、4d分析可知,Gs增加会促进叶片细胞吸收更多外界CO2而增加Ci,从而提高Pn。ET5增加能够显著提升Pn,由于抽穗开花期耗水量增加会引起叶片Gs、Tr增大的同时也利于吸收更多环境中CO2,满足光合作用所需,保持较高胞间CO2浓度以提高叶片净光合速率。不难发现,抽穗开花期耗水量对Pn影响最大,因为此时期处于需水关键期及籽粒形成关键阶段,耗水量增加会极大提升叶片光合能力以促进光合物质生产及向穗部转运与分配。
2.5 耗水过程对水稻产量的影响
作物水分生产函数反映了作物产量与水分之间的函数关系。图5显示了不同处理的水稻产量,由图可知,CK处理的产量最高为7 168.7 kg/hm2,与其他处理间均达到显著性差异(P<0.05),其次为T6处理,T2处理的产量最低,为5 612.2 kg/hm2,虽低于T1处理,但差异不显著(P>0.05)。根据试验实测的各生育时期蒸发蒸腾量,采用具有代表性的Jensen模型[16]建立水稻水分生产函数,进而得到各生育阶段的水分敏感指数,Jensen模型的表达式为
(5)
式中ya——各处理实测产量
ym——充分供水条件下的最大产量
ETmi——充分供水条件下第i阶段最大耗水量
方程拟合优度R2为0.984,且在0.01条件下达极显著水平,表明模型拟合效果较好。水分敏感指数(λ)反映了作物不同生育时期因缺水而减产的程度,水分敏感指数越高,因缺水导致的减产幅度越大。对于黑土区水稻而言,各生育时期耗水量对水稻产量影响由大到小顺序为:抽穗开花期、拔节孕穗期、分蘖中期、乳熟期、分蘖前期、分蘖后期。其中分蘖中期、拔节孕穗期、抽穗开花期λ值较高,分别为0.120、0.244、0.252,为水稻对水分亏缺的敏感时期,优先满足这些时期的供水对于水稻获得较高产量具有重要保证。

图5 各处理水稻产量Fig.5 Rice yield of each treatment
3 讨论
水稻全生育期主要阶段内的耗水强度和耗水量均大致呈现出“低-高-低”的变化趋势,在抽穗期达到最大值后逐次衰减,与石建初等[17]研究结果一致,本研究结果表明,分蘖前期耗水量及耗水强度仍然较高,其大小主要取决于插秧前土壤墒情和灌水量,因为该时段水稻幼苗叶片少且覆盖率低,田间耗水量又以棵间土壤蒸发为主,特别是表层土壤含水率的高低密切相关。研究结果还表明,分蘖期耗水模系数占全生育期43.98%,可能是由于分蘖期阶段历时最长,地上部光合产物覆盖率较低,土壤无益蒸发强度增大所致。不难发现,水稻的耗水强度在抽穗开花期达到最大,但有学者研究结果表明水稻在乳熟期蒸腾耗水强度最大[18],这可能是由于不同水稻品种、气候条件及灌溉方式所引起的。
水稻干物质的构成是光合产物在不同器官中积累与分配的结果。本研究结果表明:分蘖后期耗水量对最大叶片干物质量影响大于拔节孕穗期,这是因为分蘖后期茎蘖数基本达到高峰,耗水量增加会促进叶片的快速发育,拔节孕穗期虽然处于叶片生长的关键阶段,但其是基于分蘖后期叶片良好发育为前提条件。抽穗开花期若发生水分亏缺对茎鞘和穗干质量积累抑制作用较大,同时叶片衰老的速率加快,抑制了后期干物质的合成,导致“库源”比例失调[19],使营养物质向籽粒的转运与分配受到约束。而ET3、ET4对最大根干物质量均产生一定负效应,因为此时期适度的土壤水分调控,有利于改善土体环境的通透性,促进好气性细菌的繁殖,限制根层土壤中有毒有害物质的产生,加快有机质的分解而提高肥力、促进根系发育[20],同时对稻株生理起到先抑后促,对碳氮代谢起到削氮促碳的作用,利于氮代谢旺盛逐渐转向碳代谢,达到抑氮增糖,强化根系的作用[21],为根干物质量的积累打下重要基础。然而过于追求水稻根部干物质积累,在一定程度也抑制叶片和茎鞘营养物质向穗部转化,对于穗部籽粒的生长可能是一种负影响,这与吴岳轩等[22]研究相吻合。抽穗开花期水稻根部活性能够促进土壤水分及有机物从根系转运至地上部冠层,利于延缓叶片衰老,对于激发作物生产潜能提供必要保证[23],对最大根系伤流量研究结果表明,由于分蘖前期根系吸收水分及养分的能力较弱,耗水量对抽穗开花期根伤流量的影响并不显著;ET2对其调控效应最大,此时期供水充足会使植株幼根的伸长生长区和根毛区发育良好,有助于改善水稻根系后期的抗逆能力进而提高根伤流量[24];分蘖后期至抽穗开花期适当水分亏缺均利于提高根系活力,进而增加根系伤流量,这与汪妮娜等[23]研究结果相似。表明水分胁迫并非完全是负效应,特定发育阶段、一定水分胁迫对作物生长生理特性是有益的。本研究还发现,ET5减少对最大根干物质积累无显著影响,但却会对最大根伤流量提高起到一定促进作用,因为在水分亏缺的条件下,营养物质优先供应给地下部根系,因此对根系的生长和干物质积累影响不显著[25],而此时期适当水分胁迫能促进根系对水分及营养的吸收,提高根系活力,这可能与水稻根尖细胞对水分胁迫敏感性和生长过程中离子的积累有关[26]。而寻求适宜的灌水量以平衡根干物质积累、根系活力和地上部营养器官干物质积累之间的关系,提高穗干物质积累,是今后仍需探究的问题。
水分亏缺会引起气孔导度下降、CO2供应受阻,导致叶片光合能力降低,但严重亏水时叶片的光合作用会呈现不可逆性[27]。本研究结果表明,ET2增加会使水稻各项生理活动均处于旺盛水平,叶片气孔导度增大以加快与外界气体的交换,为作物叶片良好发育打下基础,进而对Gs产生一定正影响;ET4、ET5对Gs均存在显著正影响,但Gs对ET5的响应程度最为强烈,这是由于抽穗开花期为作物籽粒形成的最重要阶段,为了激发植株体内碳水化合物的合成,需要提高叶片气孔开度以增强自身的生理及代谢活动,以提高光合物质生产潜能并将光合产物及时向穗部转运与分配[28]。研究还表明,分蘖中期遭受水分胁迫,叶片的各项生理活动将会受到极大影响,进而对叶片细胞吸收、同化环境中CO2能力造成难以恢复的影响[29],严重影响Ci;ET3降低均对Ci和Pn起到一定促进作用,这是由于ET3减少会使叶片气孔导度变小来减少蒸腾损失,一方面体现了叶片为适应水分亏缺的自身调节机制,另一方面也是有效促进分蘖后期复水之后反弹补偿的产生,引起气孔增大,利于吸收环境CO2,使Ci增大并提供更多光合作用所需原料,对抽穗期Pn提升起到正向调控效果[30]。ET4、ET5对各光合特性指标均有显著和极显著正影响,表明此时期耗水量的增加对于叶片光合潜能发挥具有重要作用,但叶片Tr过大也会对植株产生一定的负面影响,作物在进行光合作用过程中,势必与周围环境发生气体交换,因此叶片水分不可避免地要顺着水势梯度丢失,但植株蒸腾作用会产生一定蒸腾拉力,其引起的吸收过程可为植株较高部分提供水分,给作物生长带来一定优势[31]。因此,寻求该时期作物最佳的需水量,不仅可以减少水分消耗,而且对作物生长有利。对于叶片净光合速率下降的原因,除了考虑气孔导度的大小,还需考虑胞间CO2浓度的变化;由图4a可知,随着气孔导度降低,胞间CO2浓度趋于平缓,这说明植物光合速率的下降,是气孔导度和胞间CO2浓度共同作用的结果,但结果和其他学者[32]研究结论不一致,在某种情况下,认为Ci降低是叶肉细胞光合活性增大乃至光合速率增高的结果,可能是由于光合指标测定时间、气候因素及作物品种差异造成的。
采用Jensen模型描述各处理产量对耗水过程的响应关系,结果显示,模型中水分敏感指数在整个生育期内呈两头小、中间大趋势,表明拔节孕穗期、抽穗开花期亏水对产量的影响最为显著,这与王克全等[33]的结论一致。但本研究中拔节孕穗期和抽穗开花期的水分敏感指数较程卫国等[34]的研究结果略小,由于本研究按照生育阶段接近等时段原则,将分蘖期细化为分蘖前、中、后期,生育阶段划分越细,水分敏感指数越小。水分敏感指数变化规律与水稻耗水规律相吻合,分蘖中期、拔节孕穗期、抽穗开花期耗水量对产量影响最大,分蘖中期是茎蘖生长的旺盛阶段,供水充足会促进有效分蘖的发生,易形成合理的高产群体。拔节孕穗期、抽穗开花期水稻地上部冠层覆盖率较高,营养物质逐渐从茎秆及叶片转移至穗部籽粒中,此两时期是产量形成的最关键时期,也是整个生育期的需水关键期,保证充足的供水对水稻产量的提升具有重要作用,同时也证明了Jensen模型的有效性和试验结果的可靠性。
4 结论
(1)在水稻各处理生育期内,耗水强度总体呈现先稳定上升、后逐步下降的趋势,分蘖前期各处理耗水强度在2.87~3.74 mm/d之间,至分蘖后期平均耗水强度逐渐增加至4.56 mm/d,各处理水稻耗水强度在抽穗开花期达到最大,为5.66 mm/d。水稻耗水模系数在整个分蘖期达到最大,平均值为43.98%,其次为拔节孕穗期,乳熟期耗水模系数最低,均值为16.02%。
(2)耗水过程对水稻各器官干物质积累的影响表明,各时期耗水量对叶片最大干物质量的影响由大到小顺序为:ET3、ET2、ET4、ET1;对茎鞘最大干物质量的影响由大到小顺序为:ET5、ET4、ET2、ET1、ET3;对穗最大干物质量的影响由大到小顺序为:ET5、ET4、ET2、ET6、ET3、ET1;对根最大干物质量的影响由大到小顺序为:ET3、ET2、ET4、ET5、ET1。
(3)耗水过程对水稻根系伤流量和叶片光合指标的影响表明,抽穗开花期伤流量对ET2的正响应最为显著;ET3、ET4、ET5均对其达显著负影响。ET2、ET4、ET5对叶片的Gs、Ci、Tr、Pn的正影响相比其他时期更为显著;ET1对Tr存在显著负效应;ET3均对Ci、Pn存在显著负效应。Gs与Ci呈开口向上二次函数关系;Tr与Gs、Pn与Ci均呈二次函数抛物线关系;Tr与Pn呈线性正相关关系。
(4)耗水过程对水稻产量的影响表明,分蘖中期、拔节孕穗期、抽穗开花期λ值较高,分别为0.120、0.244、0.252,为水稻对水分亏缺的敏感阶段,在黑土区有限灌溉水量条件下,水量优先分配到这3个阶段,是水稻获得高产的重要保证。
