鲜食糯玉米己醛和己醇的生成与LOX活性及其基因表达的相关性
2018-09-10牛丽影沈凌雁刘春菊李大婧刘春泉肖丽霞
牛丽影 沈凌雁 刘春菊 李大婧 刘春泉 肖丽霞
摘要:为探讨生鲜糯玉米汁主要挥发性成分己醛与己醇的生成与脂氧合酶(LOX)的关系,对4个籽粒发育期的鲜食糯玉米京甜紫花糯2号和苏玉糯11号生鲜玉米汁中己醛与己醇含量、脂氧合酶活性以及ZmLOX1、Zm-LOX2、ZmLOX3、ZmLOX10基因的相对表达量进行了测定和比较。结果显示己醛与己醇含量显著正相关(P<0.05),4种LOX基因的表达量之间显著正相关(P<0.05),但LOX活性与其他指标之间未表现出显著的相关关系。主成分分析( PCA)结果显示所有指标聚为3类:4种LOX基因表达量、己醛与己醇含量、LOX活性,三者之间未表现出显著的相关性关系。
關键词:鲜食糯玉米;己醛;脂氧合酶(LOX;基因表达量
中图分类号:S513.01
文献标识码: A
文章编号: 1000-4440(2018)05-1138-06
糯玉米为起源中国的玉米类型,作为鲜食玉米广受消费者喜爱。鲜食糯玉米蒸煮后散发的玉米香与类似青叶、青草的清香是其重要特征之一。在鲜食糯玉米挥发性成分与香气变化研究中发现,来自脂氧合酶(LOX)途径的己醛和己醇为生鲜玉米汁挥发性成分的主要成分,并可能与糯玉米的青香有关。LOX途径为LOX启动的具异戊二烯结构的多元不饱和脂肪酸(Polyunsaturated fatty acid,PUFA)的氧化过程,植物中多以亚油酸和亚麻酸为底物,在LOX作用下生成9-氢过氧化物(9-HPOT)和13-氢过氧化物(13-HPOT),然后由氢过氧化物裂解酶(HPL)作用生成的C6醛和C9醛,并在乙醇脱氢酶(ADH)的作用下生成相应的醇。其中C6的醛、醇、酮以及衍生的酯类被称为青叶挥发性成分(Green leaf volatile, GLV)。GLV在植物生理研究中被作为抗逆反应的标志性成分,在番茄、黄瓜或柑橘中为新鲜香气的表征成分,在豆类、速冻玉米、水稻等谷物中则与油脂氧化味等不良风味相关。
从基因水平研究LOX途径及风味调控是近年来的趋势。LOX是一个大基因家族,每种植物中都会有多个LOX同工酶,在番茄、苹果等植物中只有某特定的LOX同工酶调控GLV的生成。目前已知玉米中有13种LOX,ZmLOX1基因存在于成熟籽粒中,具有9-LOX和13-LOX的非传统型双位点特异性;ZmLOX2基因在玉米生长发育过程中表达.ZmLOX3存在于玉米植株的根部以及发育的胚中;ZmLOX10位于叶绿体中,主要调节GLV的生成。在对鲜食糯玉米LOX酶特性研究中发现,鲜食糯玉米中至少存在2种LOX同工酶,并且生鲜糯玉米汁的GLV则主要为己醛与己醇。为了在基因表达水平上探讨鲜食糯玉米中GLV生成和香气变化机制,本研究对2个品种4个发育期的鲜食糯玉米中己醛和己醇含量、LOX酶活性以及Zm-LOX1、ZmLOX2、ZmLOX3、Zm LOX10的相对基因表达量之间的相关性进行分析,为鲜食糯玉米香气的调控提供理论基础。
1 材料与方法
1.1 试验材料
糯玉米京甜紫花糯2号(JT)、苏玉糯11号(SY)均采自江苏省农业科学院六合动物科学基地试验田。2种糯玉米于授粉后14d、19d、25d、31d进行采收,分别对应于籽粒建成期、乳熟前期、乳熟后期和蜡熟期,记为JT1—4和SY1~4。采收当日手工去除苞叶,分离完整玉米籽粒,液氮速冻后真空包装,置于-80℃冰箱待用。
1.2 主要仪器和试剂
Agilent 7890A/5975C气相色谱质谱联用仪,美国Agilent公司产品;CAR/PDMS/DVB固相微萃取萃取头,美国supelco公司产品;HP-INNOWAX毛细管色谱柱,美国Agilent公司产品;荧光定量PCR仪,美国Roche公司产品。Trizol试剂盒,美国Gibco公司产品。
1.3 试验方法
1.3.1 己醛、己醇含量的测定冷冻玉米粒10g加入40ml 4℃预冷的蒸馏水,迅速研磨,用5ml水转移至均质仪进行均质后定容到50ml,取5ml于15ml顶空瓶中并加入搅拌子及1.6g NaCl,密封后40℃水浴平衡15min,插入萃取头(CAR/PDMS/DVB),使之与液面保持1.5cm的距离,加入搅拌子置于磁力搅拌器上,搅拌速度200 r/min,在40℃下萃取15min。色谱条件:色谱柱为HP-INNOWAX(30mx0.32mmx0.25μm)分析柱;升温程序为:起初温度40℃保持2 min,然后5μ/min升温至220cC,进样口温度250℃,解析2min,采用不分流进样;载气:He。质谱条件:离子源温度230℃,传输线温度为150℃,电离化方式为EI、电子能量70eV,质量扫描范围m/z 30~ 450,扫描速率为5.27scan/s,溶剂延迟l min。定量方法采用外标法,首先用色谱纯甲醇配置1mg/ml的贮备液,再稀释为0.2~200.0 mg/L的测定液,建立待测组分标准品的峰面积与对应质量浓度的标准曲线。
1.3.2 LOX活性的测定同牛丽影等的方法 。
1.3.3 实时荧光定量PCR测定LOX基因表达量使用Trizol试剂盒提取样品总RNA,用随机引物将RNA反转录成cDN。反转录反应条件为25℃10 min,42℃30 min.85℃5 min。根据基因序列设计特异性引物,通过实时荧光定量PCR方法,采用SYBR GREEN染料法,以GA PDH为内参基因,测定LOX基因表达量,以乳熟期甜玉米晶甜3号为对照。引物序列见表l。PCR反应条件为:94℃30s;94℃10s,60℃12s,72℃30s,循环45次,72℃单点检测信号。采用2-??法对结果进行相对定量分析。
1.4 数据分析
采用JMP(JMP 10.0,SAS Institute Inc.)进行单因素方差分析(ANOVA)和主成分分析(PCA)。各指标重复测定3次,结果以平均值±标准差表示。采用单因素方差分析(ANOVA) Tukey HSD进行差异显著性分析。
2 结果与分析
2.1 不同采收期两种鲜食糯玉米中己醛和己醇含量比较
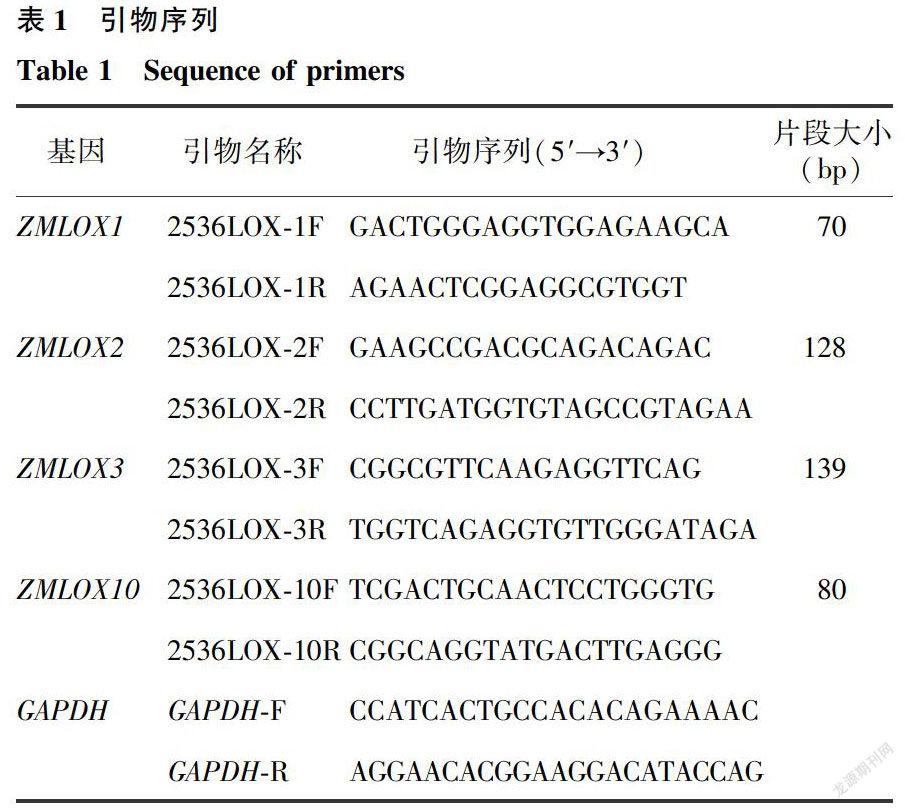
4个采收期两种鲜食糯玉米京甜紫花糯2号(JT)和苏玉糯11号(SY)中己醛含量为6.00~59.90μg/kg,己醇含量为0.11~8.29μg/kg。两种糯玉米的己醛均在籽粒建成期含量最高,在乳熟期含量最低,到蜡熟期略有回升,但蜡熟期苏玉糯11号的己醛含量与乳熟前期和乳熟后期京甜紫花糯2号差异未达显著水平(图1)。JT的己醇含量随籽粒发育期的延长而呈降低趋势,而SY的己醇含量变化不顯著。
2.2 两种鲜食糯玉米脂氧合酶(LOX)活性比较
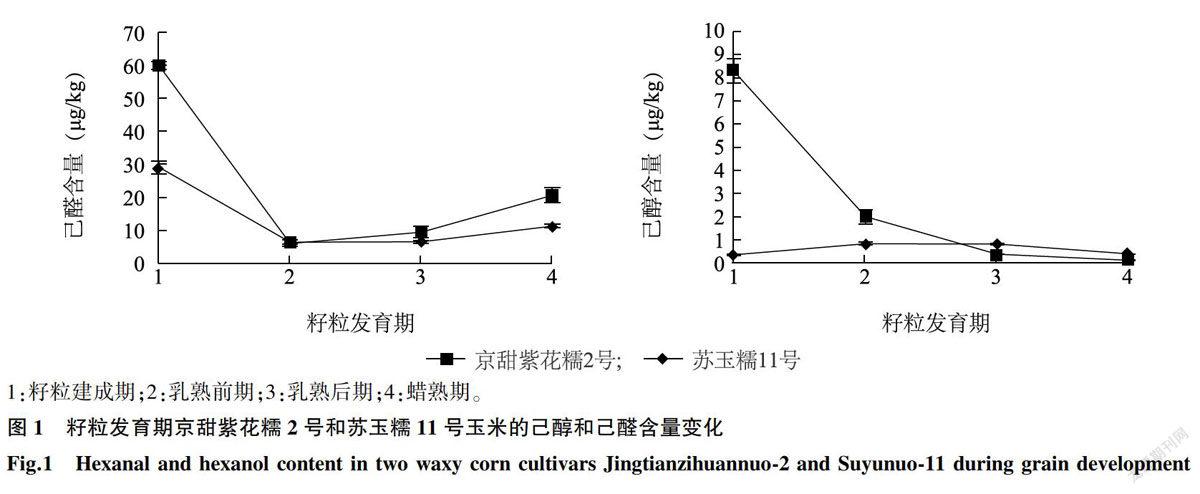
LOX活性测定结果(图2)显示,两种糯玉米京甜紫花糯2号(JT)和苏玉糯11号(SY) LOX活性均随籽粒发育期延长呈现直线上升趋势(R2=0.980 6、R2= 0.9978),在4个发育期两种玉米的LOX活性并未表现出显著差异(P>0.05)。
2.3 两种鲜食糯玉米ZmLOX1、Zm LOX2、Zm-LOX3、ZmLOX10基因表达量差异
京甜紫花糯2号的4种LOX基因表达量在发育过程中变化趋势一致,均为籽粒建成期≤乳熟后期≤乳熟前期<蜡熟期(图3)。苏玉糯11号的变化趋势与京甜紫花糯2号不同,京甜紫花糯2号的4种LOX基因表达量均为蜡熟期最高,而苏玉糯11号中ZmLOX3在乳熟后期表达量最高。另外,苏玉糯11号ZmLOX1、ZmLOX2、ZmLOX10的表达量在4个时期均等于甚至低于京甜紫花糯2号籽粒建成期的最低值。2种玉米差异较大的为乳熟前期和蜡熟期,乳熟前期京甜紫花糯2号的ZmLOX2和Zm-LOX3基因表达量分别为苏玉糯11号的117.25和4.92倍,蜡熟期京甜紫花糯2号的ZmLOX3基因表达量为苏玉糯11号的5.54倍。另外,与GLV相关的ZmLOX10基因表达量京甜紫花糯2号大于苏玉糯11号,对应4个发育期京甜紫花糯2号的基因表达量分别为苏玉糯11号的2.56、92.96、10.52和45.15倍。
2.4玉米己醇和己醛含量与LOX活性、LOX基因表达量的相关性分析
己醛和己醇是生鲜玉米汁中最主要的挥发性成分。通过调节pH以及加热等方法抑制LOX的活性可以减少二者的生成,说明LOX途径是二者生成的主要途径。相关性分析结果(表2)显示,己醛含量与己醇含量之间以及4种LOX基因表达量之间呈显著正相关关系。己醇含量与己醛含量呈显著正相关,说明生成的己醛越多,才能生成更多的己醇,但是二者的量与LOX活性、LOX基因表达量的相关性不显著。
主成分分析(PCA)结果(图4)表明,主成分l与主成分2可解释总方差的87. 0%。从图4中可以看出己醛与己醇含量、4种ZmLOX基因分别在第1象限和第2象限聚集。而样品与指标在空间上的接近则表明此样品在此指标上具有突出的特征,例如籽粒建成期的京甜紫花糯2号己醛与己醇含量最高,蜡熟期的京甜紫花糯2号ZmLOX基因表达量最高。
3 讨论
不同植物的LOX代谢机制存在差别。本研究中京甜紫花糯2号和苏玉糯11号2种糯玉米之间也存在显著差异,例如在乳熟后期京甜紫花糯2号ZmLOX1、ZmLOX2、ZmLOX10基因表达量均高于苏玉糯11号,在籽粒建成期和蜡熟期京甜紫花糯2号的己醛和己醇含量也高于苏玉糯11号。京甜紫花糯2号和苏玉糯11号糯玉米在发育过程中GLV的生成量和LOX活性存在显著变化,但是与LOX基因表达量随生长期的变化并不一致,未表现为显著的相关性关系。猕猴桃中随果实发育阶段的延长,6种LOX基因表达量不同.AdLOX1和AdLOX5在成熟期间上调,而AdLOX2、AdLOX3、AdLOX4和AdLOX6则下调。番茄中TomLOXC基因只有在成熟时才被检测到,被认为是C6醛醇生成的功能基因。ZmLOX10在玉米植株受到伤害时表达量迅速增加,促进GLV的产生。但在本研究中,在糯玉米籽粒建成期己醛与己醇含量最高,但籽粒发育后期ZmLOX10表达量最高,二者的变化并不一致。陈巧等研究发现黄瓜GLV含量与LOX基因表达间也未呈现显著相关性,而且黄瓜中Cs LOX2基因表达高峰的出现先于LOX活性高峰。但是,Zhang等发现桃成熟过程中GLV含量与PpLOX一2、PpLOX一3、PpHPL-1和PpADHl-3等基因表达水平存在一致的变化趋势。Boukobza等发现添加外源亚油酸/亚麻酸可以增加番茄GLV的生成,但是添加LOX或者乙醇脱氢酶(ADH)都不能影响GLV的生成。在氢过氧化物裂解酶(HPL)过度表达的番茄中,挥发性短链醛醇的量几乎没有改变。这些结果说明在番茄中GLV含量并不取决于LOX途径中酶的基因表达量以及酶的活性。鲜食玉米中LOX活性与C6醛醇含量之间的相关性不显著,而与亚油酸含量显著相关。谷物(如水稻、玉米)种子中LOX基因表达量与病原菌入侵引发的抗逆机制以及发芽时的生理变化有关。LOX虽然是催化脂肪氧化的酶,但是在谷物中有水的情况下会加速反应。鲜食糯玉米作为发育中的谷物种子,水分含量与果蔬接近,在蒸煮.制汁等条件下LOX作用的微环境和生化机制,及其对产品新鲜风味的影响仍待进一步研究。
