低氮诱导小麦灌浆期旗叶衰老与膜脂的关系
2018-08-10齐凌云殷俐娜王仕稳邓西平
李 倩 齐凌云 殷俐娜,,* 王仕稳, 邓西平,4
低氮诱导小麦灌浆期旗叶衰老与膜脂的关系
李 倩1,2齐凌云3,4殷俐娜1,2,3,4,*王仕稳1,2,3,4邓西平1,2,4
1中国科学院大学资源与环境学院, 北京100049;2中国科学院水利部水土保持研究所 / 黄土高原土壤侵蚀与旱地农业国家重点实验室, 陕西杨凌 712100;3西北农林科技大学资源环境学院, 陕西杨凌 712100;4西北农林科技大学水土保持研究所, 陕西杨凌 712100
小麦产量主要来自于小麦灌浆期旗叶的光合产物, 低氮造成的灌浆期旗叶早衰对小麦产量影响极大。本试验以小麦品种“长旱58”为试验材料, 在大田环境下设置低氮(120 kg hm-2)和正常氮(180 kg hm-2)处理, 研究低氮诱导的小麦旗叶衰老与膜脂的关系。结果表明, 开花14 d后, 低氮处理小麦旗叶的光合速率、叶绿素含量、旗叶总氮含量均显著降低; 旗叶中膜脂各组分含量均显著下降, DGDG/MGDG的比值升高; 以C18:3、C18:2为代表的不饱和脂肪酸含量显著下降, 以C16:0为代表的饱和脂肪酸含量显著增加, 不饱和双键指数显著降低; 此外类囊体蛋白质堆积密度也显著降低。综合分析认为低氮处理导致小麦灌浆期旗叶早衰, 早衰过程伴随着膜脂降解和组分改变, 降低了膜的流动性和通透性, 导致叶绿素降解, 使光合功能受损。同时, 植物通过调整DGDG/MGDG比例来响应低氮胁迫, 利用DGDG的双层特性来部分弥补其它双层膜脂的降解对膜功能造成的损伤。
小麦; 旗叶; 低氮; 膜脂; 衰老
小麦籽粒产量主要来自灌浆期旗叶的光合产物, 其对籽粒产量的贡献可达80%以上, 因此维持灌浆期旗叶的光合作用对小麦籽粒产量尤为重要[1]。氮素是叶绿素和色素蛋白复合体的主要成分, 也是组成蛋白质的基本元素, 由缺氮引起的灌浆期旗叶早衰是小麦减产的重要原因之一。灌浆期旗叶早衰导致叶绿素降解, 光合能力降低, 从而限制了同化物的合成和籽粒干物质的积累, 导致产量降低。植物缺氮时在显微水平表现为叶绿体数目减少, 形状变的不规则, 类囊体破裂消失, 嗜锇颗粒增多, 淀粉粒增大[2-3]; 在生化水平表现为叶绿素合成受阻, 降解速率加快, 光合碳同化酶类如Rubisco活性受到影响, PSII反应中心蛋白含量下降[4]; 在生理水平上表现为叶绿素含量和光合速率的下降[5-6]; 在形态水平表现为叶片失绿发黄, 早衰明显[7]。前人已从自由基产生和清除机制[8-9]、碳氮代谢失衡[10-11]、营养胁迫[12]、内源激素紊乱[13]等角度报道了低氮诱导早衰机制, 并且对于调控低氮诱导的叶片衰老也开展了大量研究, 包括清除自由基防止膜脂过氧化[14], 调节激素平衡[15], 调控衰老基因表达[16]等。植物叶绿体及类囊体膜在叶片光合作用中发挥重要作用, 既是光合结构的要件, 也是叶绿素蛋白、糖运输蛋白等光合作用重要蛋白的载体, 因此光合膜组成和结构的任何改变都将引起叶片结构和功能的改变[17]。在低氮诱导叶片衰老的过程中, 一个显著特征就是膜的降解和功能的丧失。
在高等植物中, 膜脂是质膜和细胞器膜的组成成分, 主要包括糖脂和磷脂, 糖脂约占膜脂总量的3/4。糖脂主要存在于叶绿体膜和类囊体膜中, 其中单半乳糖甘油二酯(monogalactosyldiacylglycerol, MGDG)和双半乳糖甘油二酯(digalactosyldiacylglycerol, DGDG)的含量分别为1/2和1/4[18]。当植物遭到逆境胁迫时, 会通过膜脂组成的变化来适应环境, 而这些变化有助于维持和恢复膜的稳定性和完整性, 从而增加植物的胁迫耐受性[19]。在低磷胁迫下, 膜中的磷脂含量降低, 为了维持膜的完整性, 会通过增加膜中的糖脂来替代缺失的磷脂[20]。在干旱胁迫下, 豇豆叶片中单半乳糖甘油二酯合酶基因、双半乳糖甘油二酯合酶基因的表达量显著升高, DGDG含量显著升高, MGDG/DGDG比值降低[21]。过表达的转基因烟草, 其MGDG和DGDG的含量增加, 植株耐盐和耐低磷能力显著提高[22-23]。在臭氧胁迫下, 菠菜叶片质体膜和类囊体膜上的糖脂明显减少[24]。以上研究结果表明, 膜脂在植物抗逆中起着重要的作用。
近年研究表明, 低氮胁迫下能维持糖脂含量和组分稳定的小麦品种具有更好的耐低氮能力[25]。小麦灌浆期旗叶衰老的过程中会伴随着叶绿体、类囊体膜及叶绿素的降解和叶片功能的丧失, 但是尚不清楚低氮胁迫下膜脂组成的变化特征, 及膜脂与低氮诱导叶片衰老的关系。本试验观测了不同氮肥水平下灌浆期小麦旗叶叶绿素含量及光合能力、含氮量、膜脂及其组分变化, 以及类囊体蛋白质的堆积密度, 旨在分析叶片衰老与膜脂的关系, 探讨低氮诱导叶片衰老的机制。
1 材料与方法
1.1 试验设计
试验地位于中国科学院长武黄土高原农业生态试验站(107°42′N, 107°40′E, 海拔1220 m)。该地年均温9.1℃, ≥10℃积温2994℃, 无霜期171 d; 年均降水量580 mm, 7月至9月降水量占全年总量的55%以上, 小麦生育期有效降雨量215.8 mm; 地下水位50~80 m。试验地土壤为中壤质黑垆土, 含有机质8.7 g kg-1、硝态氮9.8 mg kg-1、铵态氮11.1 mg kg-1、速效磷16.3 mg kg-1。
供试小麦品种为长旱58。采用完全随机区组设计, 设置正常氮(180 kg hm-2, 高产推荐用量)和低氮(120 kg hm-2) 2个氮处理, 每个处理3次重复, 小区面积为60 m2, 行距为20 cm。对所有小区基施P2O5120 kg hm-2。2015年9月28日播种, 2016年6月23日收获, 在小麦开花当天(第0天)选择开花一致的麦穗挂牌标记, 花后第14天, 取每小区30株旗叶, 一半冷冻在液氮中, 用于测定叶绿素和膜脂组成等指标, 另一半于105℃下烘30 min, 80℃下烘至恒重, 用于测定总氮含量等指标。
1.2 净光合速率和叶绿素含量的测定
于每次采样当天9:00—12:00, 用Li-6400便携式光合仪(Li-Cor, USA)测定待采叶片的净光合速率。在每个小区重复测定2个叶片, 取平均值为一个重复。每个处理3个小区即试验为3个重复。
将旗叶样品去除叶脉后剪成碎片, 称取0.2 g采用丙酮浸提比色法[26]测定叶绿素含量。在每小区重复测定2次, 取平均值为一个重复。每个处理3个小区即试验为3个重复。
1.3 叶片总氮含量测定
取不同处理条件下花后不同天数的小麦旗叶, 于105℃烘30 min, 80°C烘至恒重。采用凯氏定氮法测定叶片总氮含量。在每小区重复测定2次, 取平均值为一个重复, 每个处理3个小区即试验为3个重复。
1.4 叶片膜脂的提取、分离及脂肪酸含量测定
取不同处理条件下花后第14天的小麦旗叶, 剪碎后称取0.5 g, 按Wang和Benning[27]描述的方法提取叶片膜脂, 每处理3次重复(3个小区), 每小区1次重复。将提取的膜脂用薄层色谱(TLC)分离, 以十五烷酸(C15:0)作为内标, 采用岛津GC2014C气相色谱仪测定膜脂含量和膜脂脂肪酸组成。色谱柱为InertCap WAX (Cat. No. 1010-67162), 气化室温度250°C, 色谱柱温度140.0°C, 设置柱温箱梯度温度为: 140~160°C, 25°C min-1; 160~240°C, 8°C min-1。色谱柱流量为1.00 mL min-1, 分流比为20.0∶1。
1.5 不饱和脂肪酸双键指数的计算
根据Rawyler和Harwood等[28-29]描述的方法计算不饱和脂肪酸双键指数(double-bond index, DBI), DBI = ∑[N×P]/100, N是指每个脂肪酸分子的双键数, P为脂肪酸物质的量百分比。即
DBI = (0×P16:0+1×P16:1+2×P16:2+3×P16:3+0×P18:0+ 1×P18:1+2×P18:2+3×P18:3)/100。
1.6 统计分析
采用SPSS19.0软件统计分析数据, 用Tukey’s HSD法进行单因素方差分析, 显著性水平为0.05。用Microsoft Excel绘制柱形图。
2 结果与分析
2.1 低氮胁迫3对小麦旗叶净光合速率和叶绿素含量的影响
与开花当天相比, 花后第14天正常氮处理的旗叶净光合速率和叶绿素含量下降幅度不大, 分别为6%和2%, 而低氮处理则大幅下降30%和37% (图1), 说明低氮胁迫降低了旗叶的光合能力, 加速了花后叶绿素降解和旗叶衰老。
2.2 低氮胁迫对小麦叶片总氮含量的影响
开花当天, 低氮处理叶片总氮含量比正常氮处理低24%, 到花后第14天, 处理间差异扩大到41%; 花后第14天正常氮处理的叶片总氮含量比开花当天降低10%, 而低氮处理花后第14天的叶片总氮含量则降低30% (图2-A)。在两种氮水平下, 2个生育期的总氮/叶绿素比值没有显著差异(图2-B), 说明单位叶绿素氮含量不受氮水平的影响, 叶片总氮含量高时叶绿素含量也高, 反之亦然, 低氮胁迫会造成叶绿素含量下降。
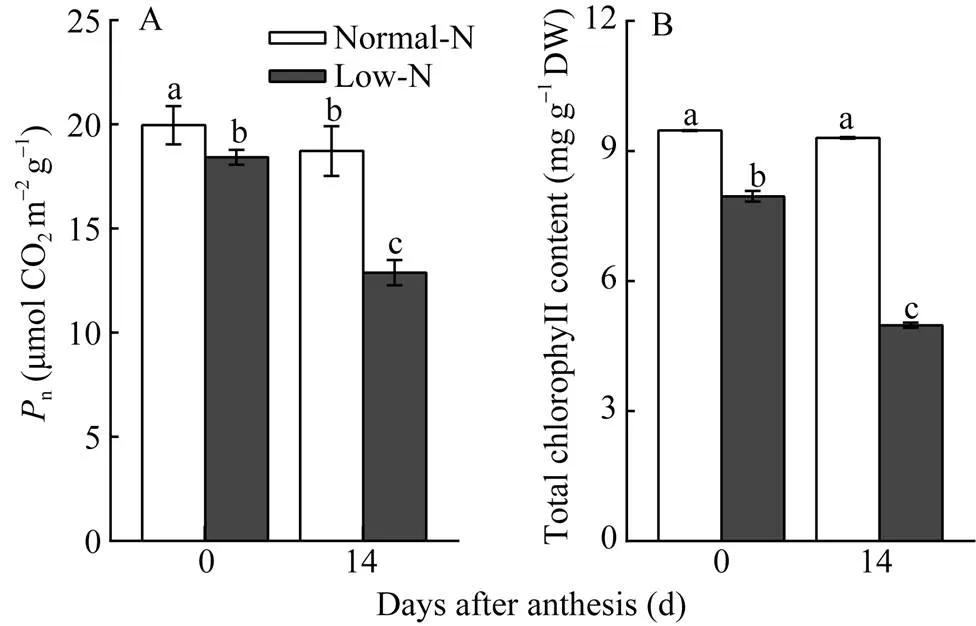
图1 低氮胁迫对小麦叶片净光合速率和叶绿素含量的影响
柱图显示平均值±标准误(= 3), 其上不同字母表示处理间差异显著(< 0.05)。
Columns represent mean ± SE (= 3) and different letters above columns mean significant difference among treatments (< 0.05).
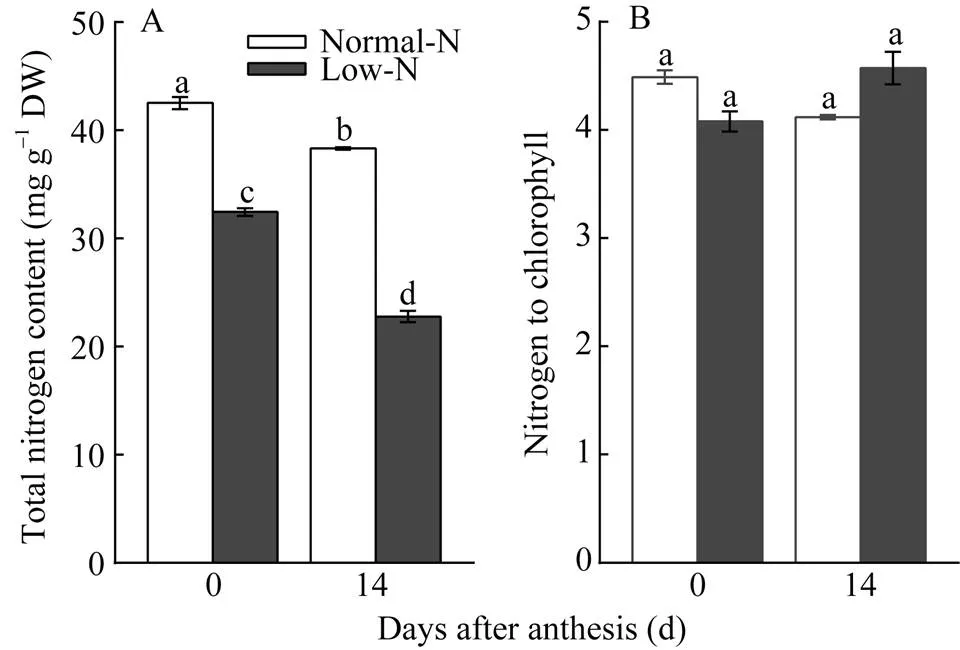
图2 低氮胁迫对小麦叶片总氮含量和总氮/叶绿素的影响
柱图显示平均值±标准误(= 3), 其上不同字母表示两处理差异显著(< 0.05)。
Columns represent mean ± SE (= 3) and different letters above columns mean significant difference among treatments (< 0.05).
2.3 低氮胁迫对小麦叶片膜脂含量和DGDG/ MGDG的影响
和正常氮处理比, 花后第14天时低氮处理的旗叶MGDG、DGDG、磷脂和总脂含量分别下降36%、32%、42%和37% (图3-A), 表明低氮胁迫对旗叶中半乳糖脂和磷脂含量有影响, 低氮可能抑制膜脂合成或者加速膜脂降解。低氮胁迫后小麦叶片中的DGDG/MGDG比值显著升高(图3-B), 说明叶片在应对逆境胁迫时通过调节MGDG和DGDG含量比例, 进而重塑膜脂来维持生物膜在逆境条件下的生理功能。
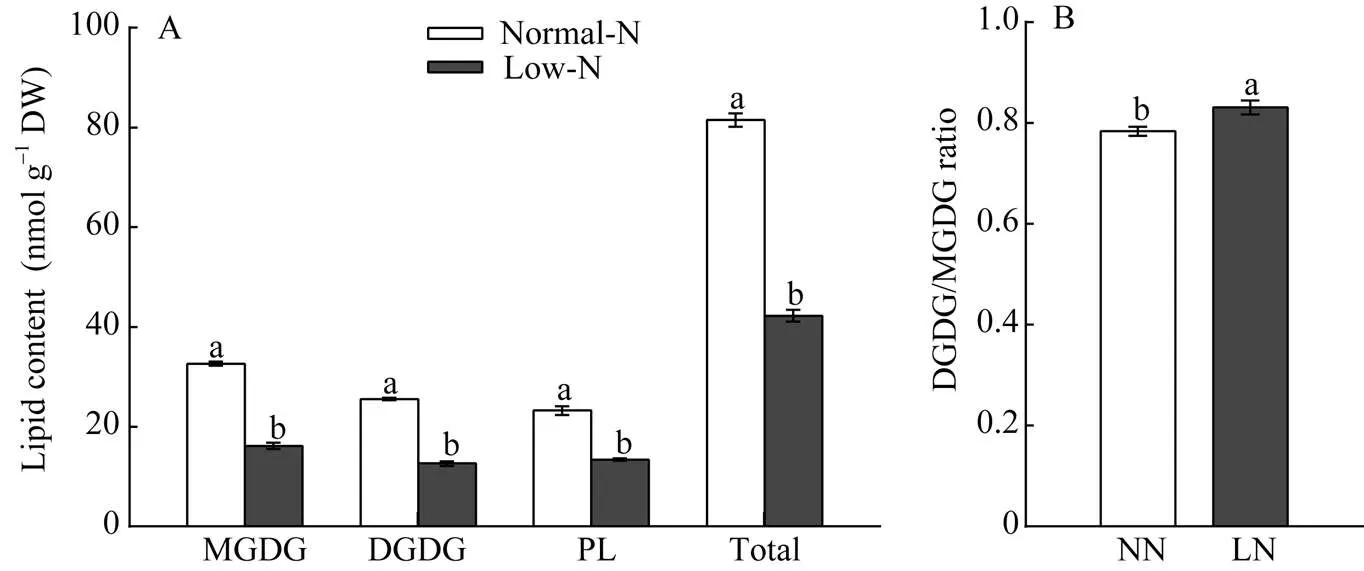
图3 低氮胁迫对小麦叶片膜脂含量(A)和DGDG/MGDG (B)的影响
MGDG: 单半乳糖甘油二酯; DGDG: 双半乳糖甘油二酯; PL: 磷脂。柱图显示平均值±标准误(= 3), 其上不同字母表示NN和LN两处理间差异显著(< 0.05)。
MGDG: monogalactosyldiacylglycerol; DGDG: digalactosyldiacylglycerol; PL: phospholipid. Columns represent mean ± SE (= 3) and different letters above columns mean significant difference between NN and LN treatments (< 0.05).
2.4 低氮胁迫对小麦叶片膜脂主要脂肪酸的影响
小麦旗叶膜脂中脂肪酸的主要成分是C18:3不饱和脂肪酸。进一步分析膜脂不同组分的脂肪酸成分, 磷脂与糖脂存在明显差异, 糖脂中C16:0、C16:1、C18:2和C18:3脂肪酸比例依次是10%、2%、3%和85%, 而磷脂中上述成分的比例分别是24%、7%、4%、12%和53%。低氮胁迫后, 小麦叶片中的MGDG、DGDG以及磷脂的不饱和脂肪酸(以C18:2和C18:3为主)含量显著下降, 饱和脂肪酸(以C16:0为主)含量显著升高(图4), 影响了膜的结构和流动性。
2.5 低氮胁迫对小麦叶片膜脂双键指数的影响
低氮胁迫后, 小麦旗叶中MGDG、PL和总脂的DBI显著下降, 但是DGDG的DBI无显著变化(图5), 说明低氮胁迫对DGDG的影响较其他种类的膜脂弱。

图4 低氮胁迫对小麦叶片中膜脂主要脂肪酸组分的影响
A: 单半乳糖甘油二酯的脂肪酸组分; B: 双半乳糖甘油二酯的脂肪酸组分; C: 磷脂的脂肪酸组分; D: 总膜脂的脂肪酸组分。柱图显示平均值±标准误(= 3), 其上不同字母表示不同脂肪酸组分间差异显著(< 0.05)。
A: fatty acid compositions of monogalactosyldiacylglycerol; DGDG: fatty acid compositions of digalactosyldiacylglycerol; PL: fatty acid compositions of phospholipid; D: fatty acid compositions of total lipids. Columns represent mean ± SE (= 3) and different letters above columns mean significant difference among fatty acid compositions (< 0.05).

图5 低氮胁迫对小麦叶片中膜脂双键指数的影响
DBI: 双键指数; MGDG: 单半乳糖甘油二酯; DGDG: 双半乳糖甘油二酯; PL: 磷脂。柱图显示平均值±标准误(= 3), 其上不同字母表示NN和LN两处理间差异显著(< 0.05)。
DBI: double-bond index; MGDG: monogalactosyldiacylglycerol; DGDG: digalactosyldiacylglycerol; PL: phospholipid. Columns represent mean ± SE (= 3) and different letters above columns mean significant difference between the NN and LN treatments (< 0.05).
2.6 低氮胁迫对小麦叶片类囊体蛋白质堆积密度的影响
低氮胁迫后小麦叶片中MGDG/叶绿素、DGDG/叶绿素、PL/叶绿素以及总脂/叶绿素的比值均显著升高(图6)。类囊体蛋白质堆积密度的显著降低, 影响了小麦旗叶的光合系统、运输系统等结构, 从而使类囊体的生理功能受到严重破坏。
3 讨论
叶绿素含量既是衡量叶片光合能力的常用参数, 也是叶片衰老的重要监测指标。氮素水平与小麦生育后期旗叶衰老有密切关系, 低氮处理叶片比正常氮处理叶片更早衰老, 并且后期的衰老进程也更快[30]。本试验观察到花后第14天, 低氮处理旗叶的叶绿素含量和净光合速率均显著下降, 且旗叶明显失绿发黄。说明旗叶早衰。本试验氮处理后旗叶总氮含量显著降低, 但是总氮/叶绿素值无显著差异, 这与前人试验结果[31-33]一致, 说明低氮胁迫促使小麦灌浆期旗叶早衰, 加重了叶片的衰老。
低氮处理后拟南芥角果在衰老过程中膜脂发生了降解, 并将降解产物转移到种子中用于储藏脂三酰甘油的合成, 说明低氮胁迫导致膜脂含量及组分变化[34]。MGDG与DGDG在结构上有较大差异, DGDG为双层膜脂, MGDG为非双层膜脂, 且MGDG为DGDG的合成底物[35]。膜脂中MGDG和DGDG的含量以及DGDG/MGDG的比例会影响生物膜的结构和功能。Gasulla等[36]比较了耐旱植物(和) 和干旱敏感植物(和拟南芥)在干燥及复水过程中膜脂组成的变化, 以及膜脂合成基因表达情况, 发现干旱胁迫下耐旱植物中总脂含量维持稳定, 但是膜脂的组成发生了较大的变化, 最主要的变化是约50%的MGDG水解, 其中一部分水解成DAG用来合成磷脂, 另一部分则通过DGD1/DGD2途径合成DGDG, 随后通过SFR2形成寡聚半乳糖酯, 这些合成的磷脂和DGDG以及寡聚半乳糖酯均用于稳定质膜及亚细胞器膜。本研究表明, 低氮处理导致的早衰叶片中MGDG、DGDG、PL和总脂的含量均显著下降, 但是DGDG/MGDG的比值却显著升高, 这是因为低氮胁迫促使膜脂降解, 但是作为DGDG的合成底物, MGDG部分转化为DGDG, 而DGDG的双分子层结构会部分补偿磷脂双分子层的损伤, 对膜进行重塑, 有助于在最大程度上维持膜稳定。同时膜脂脂肪酸比例也发生了变化, 其中不饱和脂肪酸(C18:2和C18:3)含量显著下降, 而饱和脂肪酸(C16:0)含量显著上升, 反映出膜的流动性受到损伤。分析膜脂DBI, 发现低氮胁迫下MGDG、DGDG、磷脂以及总脂的DBI均显著下降, 说明膜的流动性和通透性受到破坏。以上结果表明, 低氮胁迫导致小麦旗叶中膜脂降解, 膜的流动性和通透性受到损伤, 影响了小麦旗叶的正常生理功能。
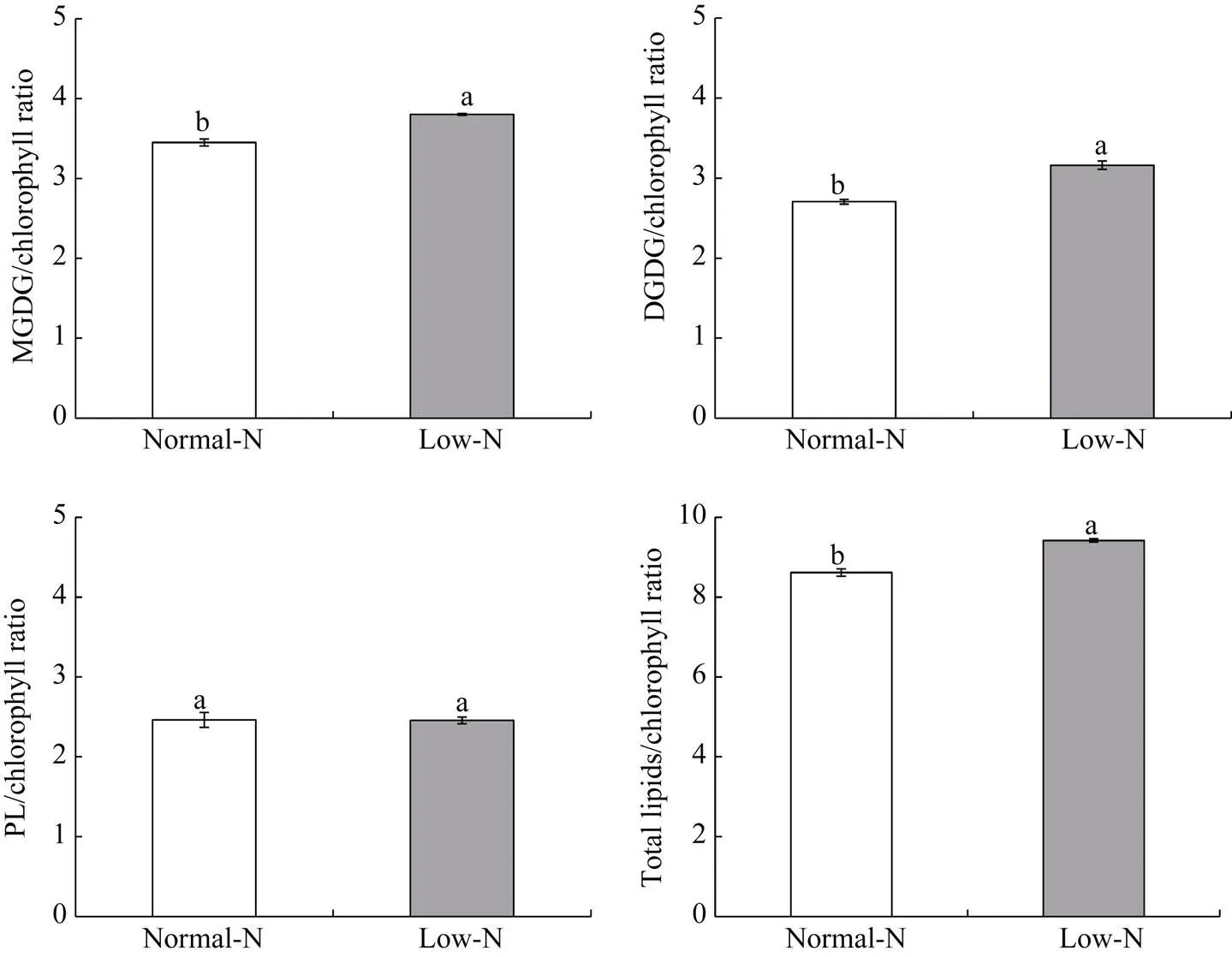
图6 低氮胁迫对小麦叶片中膜脂与叶绿素含量比值的影响
MGDG: 单半乳糖甘油二酯; DGDG: 双半乳糖甘油二酯; PL: 磷脂。柱图显示平均值±标准误(= 3), 其上不同字母表示NN和LN两处理间差异显著(< 0.05)。
MGDG: monogalactosyldiacylglycerol; DGDG: digalactosyldiacylglycerol; PL: phospholipid. Columns represent mean ± SE (= 3) and different letters above columns mean significant difference between NN and LN treatments (< 0.05).
类囊体是植物光合作用的主要场所, 几乎所有的光合蛋白复合体均与叶绿素结合在类囊体膜上。类囊体膜主要由糖脂(MGDG和DGDG)组成, 因此, 糖脂/叶绿素的比值很好地反映出类囊体上光合蛋白的堆叠密度[37]。糖脂与叶绿素的比值越高表明蛋白质的堆积密度越低[38]。胡锋等[39]研究发现, 低氮胁迫时拟南芥光合能力下降的同时伴随着MGDG/叶绿素和DGDG/叶绿素的显著增加, 表明缺氮时类囊体中光合蛋白的堆积密度有下降的趋势。在本试验中, 低氮处理旗叶花后第14天MGDG/叶绿素比值、DGDG/叶绿素比值以及总脂/叶绿素比值显著增加, 说明低氮胁迫降低了小麦旗叶类囊体中光合蛋白的堆积密度。这不仅影响类囊体的结构, 而且影响光合蛋白复合体的生理功能, 从而使光合作用受到负面影响, 进一步加速了叶片的衰老。
4 结论
低氮胁迫下小麦灌浆期旗叶衰老伴随着膜脂含量的降低和不饱和脂肪酸双键指数降低, 膜脂降解, 膜的流动性和通透性遭到破坏; 与此同时蛋白质堆积密度降低, 叶绿素含量降低, 导致类囊体膜的结构和功能受损, 使叶片的光合功能破坏, 引起早衰。MGDG向DGDG的转化是生物膜应对低氮胁迫的响应表现之一, 通过DGDG的双层结构部分补偿膜的损伤, 维持膜的结构和功能。
[1] 李世清, 王瑞军, 张兴昌, 伍维模, 邵明安. 小麦氮素营养与籽粒灌浆期氮素转移的研究进展. 水土保持学报, 2001, 18: 106–111 Li S Q, Wang R J, Zhang X C, Wu W M, Shao M A. Research advancement of wheat nitrogen nutrition and nitrogen transportation in wheat grain filling., 2001, 18: 106–111 (in Chinese with English abstract)
[2] Hashimoto H, Kura Hotta M, Katoh S. Changes in protein content and in the structure and number of Chloroplasts during leaf senescence in rice seedlings., 1989, 30: 705–707
[3] Barton R. The production and behavior of phytoferritin particles during senescence of phasolusleave., 1970, 94: 73–77
[4] Kolber Z, Falkowski P G. Effects of growth irradiance and nitrogen limitation on photosynthetic energy conversion photosystem II., 1988, 88: 923–929
[5] 孙虎. 氮肥对番茄衰老调控及产量的影响. 北方园艺, 2016, 40(24): 35–37 Sun H. Effect of nitrogen fertilizer on tomato aging control and production., 2016, 40(24): 35–37 (in Chinese with English abstract)
[6] 曾建敏, 崔克辉, 黄见良, 贺帆, 彭少兵. 水稻生理生化特性对氮肥的反应及与氮利用效率的关系. 作物学报, 2007, 33: 1168–1176 Zeng J M, Cui K H, Huang J L,He F, Peng S B. Responses of physio-biochemical properties to N-fertilizer application and its relationship with nitrogen use efficiency in rice (L.)., 2007, 33: 1168–1176 (in Chinese with English abstract)
[7] 张绪成, 于显枫, 高世铭. 氮素对高大气CO2浓度下小麦叶片光合功能的影响. 作物学报, 2010, 36: 1362–1370 Zhang X C, Yu X F,Gao S M. Effects of nitrogen nutrition on photosynthetic function of wheat leaves under elevated atmospheric CO2concentration., 2010, 36: 1362–1370 (in Chinese with English abstract)
[8] Halliwell B. Chloroplast Metablism: The Structure and Function of Chloroplasts in Green Leaf Cells. Oxford: Charenden Press, 1981
[9] 许长成, 邹琦. 大豆叶片早促衰老及其与膜质过氧化的关系. 作物学报, 1993, 19: 359–364 Xu C C, Zou Q. The acceleration of senescence of soybean leaves induced by drought and its relation to membrane lipid peroxidation., 1993, 19: 359–364 (in Chinese with English abstract)
[10] Chen D, Wang S, Xiong B, Deng X. Carbon/nitrogen imbalance associated with drought-induced leaf senescence in sorghum bicolor., 2015, 10: e0137026
[11] 曹蓓蓓, 王仕稳, 齐凌云, 陈道钳, 殷俐娜, 邓西平. 小麦苗期叶片碳氮平衡与低氮诱导的叶片衰老的关系. 麦类作物学报, 2017, 37: 1–7 Cao B B, Wang S W, Qi L Y, Chen D Q, Yin L N, Deng X P. Carbon/nitrogen balance involved in nitrogen deficiency induced leaf senescence in wheat seedling., 2017, 37: 1–7 (in Chinese with English abstract)
[12] Sinclair T R, de Wit C T. Comparative analysis of photosynthate and nitrogen requirements in the production of seeds by various crops., 1975, 189: 565–567
[13] Wanda Y, Miura K, Wantanabe K. Effect of source to sink ratio on carbohydrate production and senescence of rice flag leaves during the ripening period., 1993, 62: 547–553
[14] 李淑文, 文宏达, 薛宝民, 齐永清, 肖凯. 小麦高效吸收利用氮素的生理生化特性研究进展, 麦类作物学报, 2003, 23(4): 131–135 Li S W, Wen H D, Xue B M,Qi Y Q, Xiao K. Advances on the physiological and biochemical characteristics of high nitrogen use efficiency in wheat., 2003, 23(4): 131–135 (in Chinese with English abstract)
[15] 赵平, 林克惠, 郑毅. 氮钾营养对烟叶衰老过程中内源激素与叶绿素含量的影响. 植物营养与肥料学报, 2005, 11: 379–384 Zhao P, Lin K H, Zheng Y. Effect of N and K nutrition on chlorophyll content and endogenous hormones in the process of tobacco senescence., 2005, 11: 379–384 (in Chinese with English abstract)
[16] 王汉忠, 赵福庚, 张国珍. 多胺延缓植物衰老的机制. 山东农业大学学报, 1995, 26: 227–232 Wang H Z, Zhao F G, Zhang G Z. Retardation of plant senescence by polyamines., 1995, 26: 227–232 (in Chinese with English abstract)
[17] Zhang M, Deng X, Yin L, Qi L, Wang X, Wang S, Li H. Regulation of galactolipid biosynthesis by overexpression of the ricegene contributes to enhanced aluminum tolerance in tobacco., 2016, 7: 337
[18] Block M A, Dorne A J, Joyard J, Douce R. Preparation and characterization of membrane-fractions enriched in outer and inner envelope membranes from spinach-chloroplasts II biochemical-characterization., 1983, 258: 13281–13286
[19] Ohlrogge J, Browse J. Lipid biosynthesis., 1995, 7: 957–970
[20] Harwood J L. Recent advances in the biosynthesis of plant fatty acids., 1996, 1301: 7–56
[21] Shimojima M, Watanabe T, Madoka Y, Koizumi R, Masuda K, Yamada K, Masuda S, Ohta H. Differential regulation of two types of monogalactosyldiacylglycerol synthase in membrane lipid remodeling under phosphate-limited conditions in sesame plants., 2013, 469: 1–10
[22] Wang S, Uddin M I, Tanaka K, Yin L, Shi Z, Qi Y, Mano J, Matsui K, Shimomura N, Sakaki T, Deng X, Zhang S. Maintenance of chloroplast structure and function by overexpression of the ricegene leads to enhanced salt tolerance in tobacco., 2014, 165: 1144–1155
[23] 史中惠, 王仕稳, 殷俐娜, 张梅娟, 邓西平. 超表达水稻MGD基因()烟草植株的耐低磷胁迫能力. 西北农林科技大学学报(自然科学版) . 2013, 41(10): 97–104 Shi Z H, Wang S W, Yin L N,Zhang M J, Deng X P. Tolerance of tobacco plants withgene to low phosphorus stress.(Nat Sci Edn), 2013, 41(10): 97–104 (in Chinese with English abstract)
[24] Sakaki T, Saito K, Kawaguchi A, Kondo N, Yamada M. Conversion of monogalactosyldiacylglycerols to triacylglycerols in ozone-fumigated spinach leaves.,1990, 94: 766–772
[25] 齐凌云, 张梅娟, 曹蓓蓓, 殷俐娜, 王仕稳, 邓西平. 小麦苗期叶片膜质组成对低氮胁迫的响应及其与耐低氮的关系. 植物生理学报, 2017, 53: 1039–1050 Qi L Y, Zhang M J, Cao B B,Yin L N, Wang S W, Deng X P. Effect of nitrogen deficiency on leaf lipid composition in different nitrogen deficiency tolerant wheat () cultivars during seedling stage., 2017, 53: 1039–1050 (in Chinese with English abstract)
[26] 高俊凤. 植物生理学试验指导. 北京: 高等教育出版社, 2006. pp 74–77 Gao J F. Experimental Guidance for Plant Physiology. Beijing: Higher Education Press, 2006. pp 74–77 (in Chinese)
[27] Wang Z, Benning C.polar glycerolipid profiling by thin layer chromatography (TLC) coupled with gas-liquid chromatography (GLC)., 2011, 49: e2518
[28] Rawyler A, Pavelic D, Gianinazzi C, Oberson J, Braendle R. Membrane lipid integrity relies on a threshold of ATP production rate in potato cell cultures submitted to anoxia., 1999, 120: 293–300
[29] Harwood J L. Fatty acid metabolism., 1988, 39: 101–138
[30] 冯波, 王法宏, 刘延忠, 孔令安, 张宾, 李升东, 司纪升. 施氮水平对不同栽培模式小麦旗叶衰老及产量的影响. 中国农学通报, 2010, 26(8): 189–193 Feng B, Wang F H, Liu Y Z,Kong L A, Zhang B, Li S D, Si J S. Effect of nitrogen application on senescence of flag leaf and grain yield in wheat in different planting models., 2010, 26(8): 189–193 (in Chinese with English abstract)
[31] 许强, 王彦才, 马宏玮. 宁夏春小麦缺氮导致减产的生理机理研究. 干旱地区农业研究, 1999, 17(3): 56–61 Xu Q, Wang Y C, Ma H W. Study on physiological mechanism of the decline in spring wheat production caused by inscofficient nitrogen in Ningxia., 1999, 17(3): 56–61 (in Chinese with English abstract)
[32] 何萍, 金继运, 林葆. 氮肥用量对春玉米叶片衰老的影响及其机理研究. 中国农业科学, 1998, 31(3): 66–71 He P, Jin J Y, Lin B. Effect of n application rates on leaf senescence and its mechanism in spring maize., 1998, 31(3): 66–71 (in Chinese with English abstract)
[33] 王亚江, 魏海燕, 颜希亭, 葛梦婕, 孟天瑶, 张洪程, 戴其根, 霍中洋, 许轲, 费新茹. 光、氮及其互作对超级粳稻产量和氮、磷、钾吸收的影响. 作物学报, 2014, 40: 1235–1244 Wang Y J, Wei H Y , Yan X T, Ge M J, Meng T Y, Zhang H C, Dai Q G, Huo Z Y, Xu K, Fei X R. Effects of light, nitrogen and their interaction on grain yield and nitrogen, phosphorus and potassium absorption insuper rice., 2014, 40: 1235–1244 (in Chinese with English abstract)
[34] 禹晓梅, 王荣, 李唯奇. 拟南芥角果衰老过程中膜脂的变化. 植物分类与资源学报, 2014, 36: 177–186 Yu X M, Wang R, Li W Q. Changes in membrane lipids during silique senescence in Arabidopsis., 2014, 36: 177–186 (in Chinese with English abstract)
[35] Douce R, Joyard J. Lipids: structure and function. New York: Academic Press, 1980. pp 221–225
[36] Gasulla F, vom Dorp K, Dombrink I, Dormann P, Bartels D. The role of lipid metabolism in the acquisition of desiccation tolerance in: a comparative approach., 2013, 75: 726–741
[37] Kirchhoff H, Sharpe R M, Herbstova M, Yarbrough R, Edwards G E. Differential mobility of pigment-protein complexes in granal and agranal thylakoid membranes of C3and C4plants., 2013, 161: 497–507
[38] Haferkamp S, Kirchhoff H. Significance of molecular crowding in grana membranes of higher plants for light harvesting by photosystem II., 2008, 95:129–134
[39] 胡锋, 黄俊丽, 秦峰, 岳彩黎, 王贵学. 植物叶绿体类囊体膜及膜蛋白研究进展. 生命科学, 2011, 23: 291–198Hu F, Huang J L, Qin F,Yue C L, Wang G XProgress in chloroplast thylakoid membrane and membrane proteins., 2011, 23: 291–198 (in Chinese with English abstract)
Relationship between Lipid and Flag Leaf Senescence Induced by Low Nitrogen Stress during Grain Filling of Wheat
LI Qian1,2, QI Ling-Yun3,4, YIN Li-Na1,2,3,4,*, WANG Shi-Wen1,2,3,4, and DENG Xi-Ping1,2,4
1College of Natural Resources and Environment, University of Chinese Academy of Sciences, Beijing 100049, China;2State Key Laboratory of Soil Erosion and Dryland Farming on the Loess Plateau / Institute of Soil and Water Conservation, Chinese Academy of Sciences and Ministry of Water Resource, Yangling 712100, Shaanxi, China;3Institute of Soil and Water Conservation, College of Life Science, Northwest A&F University, Yangling 712100, Shaanxi, China;4College of Natural Resources and Environment, Northwest A&F University, Yangling 712100, Shaanxi, China
Nitrogen (N) deficiency induces flag leaf senescence at the grain-filling stage of wheat resulting in yield decrease. In this study, we carried out a field experiment to investigate the relationship between N-deficiency-induced flag leaf senescence and lipid change using wheat variety ‘Changhan 58’. Under the low-N (120 kg ha–1) condition, the photosynthetic rate, chlorophyll content, total N content, and contents of all lipid components of flag leaf at 14 days after flowering were significantly lower than those under normal-N (180 kg ha–1) condition, while the DGDG/MGDG ratio in low-N treatment increased. The content of unsaturated fatty acids, such as C18:3 and C18:2 decreased significantly, whereas the contents of saturated fatty acids, such as C16:0, increased significantly, leading to the reduction of index of unsaturated double bounds. In addition, thylakoid protein packing density also decreased under N-deficiency condition. These results indicate that N-deficiency-induced leaf senescence is characterized with lipid degradation and components changes, which result in negative effects on reductions of membrane permeability, chlorophyll content and photosynthetic rate. Simultaneously, the DGDG/MGDG ratio of plants is adjusted in response to N deficiency to alleviate damages of membrane function caused bby lipid degradation.
wheat; flag leaf; nitrogen deficiency; membrane lipids; senescence
本研究由国家自然科学基金项目(31200206), 国家科技支撑计划项目(2015BAD22B01), 陕西省科技新星项目(2016KJXX-66)和中国科学院青年创新促进会项目(2015389)资助。
This study was funded by the National Natural Science Foundation of China (31200206), the National Key Technology R&D Program of China (2015BAD22B01), the Program for Scientific and Technological Star of Shaanxi Province (2016KJXX-66), and the Youth Innovation Promotion Association of the Chinese Academy of Sciences (2015389).
URL: http://kns.cnki.net/kcms/detail/11.1809.S.20180511.1037.002.html
2018-04-11;
2018-05-14.
10.3724/SP.J.1006.2018.01221
殷俐娜, E-mail:linayin@nwsuaf.edu.cn
E-mail: liqian515@mails.ucas.edu.cn
2017-11-09;
