The biotypes and host shifts of cotton-melon aphids Aphis gossypii in northern China
2018-08-06ZHANGShuaiLUOJunyuWANGLiWANGChunyiLiminZHANGLijuanZHUXiangzhenCUIJinjie
ZHANG Shuai, LUO Jun-yu, WANG Li, WANG Chun-yi, LÜ Li-min, ZHANG Li-juan, ZHU Xiang-zhen,CUI Jin-jie
State Key Laboratory of Cotton Biology, Institute of Cotton Research, Chinese Academy of Agricultural Sciences, Anyang 455000,P.R.China
Abstract Aphis gossypii is a globally distributed species and therefore has a highly variable life cycle. Populations of A. gossypii in northern China exhibit greater genotypic diversity and a broader host range, yet the details of life cycles of different biotypes is stillunclear. In this study, the Cytb and 16S gene regions of A. gossypii collected from 5 common summer hosts and 4 primary hosts were analyzed. A totalof 57 haplotypes were obtained from 1 046 individuala. gossypii sequences. The sequence included 44 variable sites, 27 of which were parsimony informative sites and 17 of which were singleton variable sites. The most frequent 3 haplotypes were found in 896 individuals, representing a totalof 85.7% of all individuals and 36 haplotypes were found in 1 individual. A neighbor-joining tree was constructed using 21 haplotypes that were found in more than 2 individuals. Considering the individual host plant, 5 biotypes were identified. Type 1 corresponded exactly to the cucurbit host-race and the other 4 biotypes were found as cotton host-races. Type 3 was the most abundant biotype in cottonfields in northern China.
Keywords: cotton-melon aphid, Cytb and 16S genes, haplotype, biotype, cottonfield
1. Introduction
The cotton-melon aphid, Aphis gossypii Glover, is a mosaic of sympatric host-races (Carletto et al. 2009b).This pest colonizes more than 600 plant species, including many important crops such as cotton, cucurbits, citrus,aubergine, potato, and okra (Blackman and Eastop 2000).Economic losses caused by this aphid result from its direct physical damage due to extraction of carbohydrates and amino acids from the plant phloem, and also due to spreading of plant viral diseases (Chan et al. 1991). In northern China, the species is an important insect pest of cotton and has breakouts 2 times each year during the cotton seedling stage and during summer (Li et al. 2013).A. gossypii that specializes in cucurbits is by far the most serious pest of Cucurbitaceae crops worldwide (Thomas et al. 2016).
A. gossypii is a globally distributed species and thus has a highly variable life cycle. It has been described as holocyclic in China, Japan, Korea, India, and the USA, where there are very cold winters (Margaritopoulos et al. 2007). In most places where it is found, including in Europe and Africa, it reproduces continuously by apomictic parthenogenesis(Blackman and Eastop 2000). Different genetic diversity has been observed in A. gossypii (Vanlerberghe-Masutti et al.1999; Brevault et al. 2008; Charaabi et al. 2008; Carletto et al. 2009b; Razmjou et al. 2010; Komazaki et al. 2011).
Genetic diversity is correlated with host type, and the existence of A. gossypii biotypes was determined by host transference experiments (Carletto et al. 2009b). Moreover,RAPD bands can be considered as diagnostic loci since they werefixed in A. gossypii populations collected on cucurbits and absent in those collected on noncucurbit host plants (Vanlerberghe-Masutti and Chavigny 1998). Using A. gossypii microsatellite loci (Vanlerberghe-Masutti et al.1999), Carletto et al. (2009b) found that genotypes were associated with host plants, and 5 host-races including cucurbits, cotton, eggplant, potato, and chili or sweet pepper were identified at a large geographical scale. We found that 31 A. gossypii populations in northern China could be classified as 3 host biotypes (Luo et al. 2016). Mitochondrial genomes have been frequently used to explore insect biotype (Frohlich et al. 1999); a previous study found that there was high divergence in cytochrome b sequences between cucurbit and cotton host-races of A. gossypii(Carletto et al. 2009a). Based on complete mitochondrial sequences, we developed a molecular marker with 5 singlenucleotide polymorphisms to distinguish the biotypes (Wang et al. 2016).
Previous research has demonstrated that A. gossypii can be holocyclic in northern China. Generally, A. gossypii eggs hatch in March on primary hosts, then alate adults appear and migrate to cottonfields, where seedlings emerge in late April to mid-May. In autumn, alate adults return to the primary hosts to mate and oviposit (Xia 1997). More than 10 of these primary hosts are extremely abundant in China, including hibiscus (Hibiscus syriacus), Chinese prickly ash (Zanthoxylum bungeanum), and pomegranate(Punica granatum) (Ebert and Cartwright 1997; Xia 1997;Wu et al. 2004).
Populations of A. gossypii in northern China exhibit greater genotypic diversity and broad host range, but the details of life cycles of different biotypes remain unclear.The objective of the current study was to determine the host range of different host biotypes and the life cycle. To achieve this goal, A. gossypii collected from northern China in 2014 were analyzed using a portion of the mitochondrial gene.
2. Materials and methods
2.1. Plants and insects
In 2014, 5 common summer host species (cotton,cucumber, zucchini, muskmelon, and kidney bean) were planted in afield in the same site in Anyang (36°5´34.8´´N,114°31´47.19´´E), Henan Province, China, samples were collected during different seasons from those summer hosts (Wang et al. 2016). Populations from the urban ornamental plants (hibiscus, pomegranate, Chinese prickly ash, and Sophora japonica) in the village of Anyang were also collected, and combined with collected data previously reported (Wang et al. 2016).
Samples of wingless A. gossypii aphids were collected from cottonfields in northern China in late August 2014 at a totalof 20 sites, including 6 populations in Hebei Province,6 in Henan Province (including Anyang), and 8 in Shandong Province, as done previously reported (Luo et al. 2016).Only 1 individual per plant from 8-48 cotton plants per site was collected to avoid sampling the offspring of a single female.
2.2. Sample sequencing
Genomic DNA from single individuals was extracted using a TIANamp Genomic DNA Kit (Tiangen, Beijing, China)following the protocol described by the manufacturer, and all extractions were stored at −20°C. Cytb and 16S gene regions were amplified with CytbF and 16SR primers (Wang et al. 2016; Zhang et al. 2016). The amplified products were sequenced at Shanghai Sangon Biotech, following trimming of the fragment to remove DNA data with poor quality.
2.3. Data analyses
The number of polymorphic sites, haplotype diversity,nucleotide diversity, average number of nucleotide differences, and the number of haplotypes were calculated using DnaSP ver. 5.0 (Librado and Rozas 2009). Samples with haplotypes representing more than 2 individuals were analyzed. The alignment of the haplotype sequences was done manually in MEGA ver. 6.0 (Tamura et al. 2013),and 1 000 neighbor-joining (NJ) bootstrap replicates were performed. Then a NJ tree was constructed, and built using the iTOL webpage (http://itol.embl.de/) (Letunic and Bork 2016). Venny ver. 2.0 (http://bioinfogp.cnb.csic.es/tools/venny) was used to identify overlapping (and unique)haplotypes and generate Venn diagrams.
3. Results
3.1. ldentification of haplotypes
All sequences were trimmed with ContigExpress Software,and sequences with lengths of 577 nt were kept. A totalof 1 046 individuala. gossypii sequences were selected.Using DnaSP Software, 57 haplotypes were obtained from 1 046 individuala. gossypii sequences. All sequences were unambiguously aligned and no insertions or deletions were found. The sequence covered 46 variable sites, 27 of which were parsimony informative sites, and 17 of which were singleton variable sites (Fig. 1).
The 3 most frequent haplotypes were Hap1, found in 568 individuals, Hap4, found in 212 individuals, and Hap 3, found in 116 individuals. Those top 3 most frequent haplotypes were found in 896 individuals, which represented 85.7%of all individuals. There were 36 haplotypes that were found in only 1 individual, which amounted to 63.2% of all haplotypes (Fig. 2).
3.2. Haplotypes in different host plants
The haplotypes of 9 host plants were analyzed. The number of haplotypes (Nh) of each host was significantly variable; the average Nh over all 9 populations was 10.44, ranging from 2 to 21, which were in populations on S. japonica and zucchini, respectively. The average nucleotide diversity found over all 9 populations was 0.263%, ranging from 0.084 to 0.385%, which were in populations on pomegranate and kidney bean, respectively.The haplotype diversity (Hd) ranged from 0.095 to 0.800.The highest haplotype diversity value was observed in populations on Chinese prickly ash. The populations of cucumber and zucchini also showed the highest haplotype diversity with 0.795 and 0.737, respectively. The lowest haplotype diversity values were observed in populations on S. japonica; populations on hibiscus and muskmelon also had a lower diversity with values of 0.155 and 0.401,respectively (Table 1).
In this study, the number of haplotypes on 3 aphid populations from primary hosts of hibiscus, pomegranate,and Chinese prickly ash were 77, 4, and 20, respectively.The wingless aphid populations on hibiscus collected in November contained all 3 haplotypes. The winged aphid populations on pomegranate collected in April contained all 4 haplotypes (Hap1, Hap4, Hap10, Hap11). The aphid populations on Chinese prickly ash collected at 3 times had 1 common haplotype (Hap1). Wingless aphid populations in November contained more haplotypes (9 unique haplotypes)than the other 2 sampling times of wingless (in April; 3 haplotypes) and winged (in November; 4 haplotypes) aphids(Fig. 3-A). Four aphid populations on zucchini collected at 3 times had 4 common haplotypes (Hap1, Hap3, Hap4,Hap16). Winged aphid populations in May contained fewer haplotypes than at the other 3 times, which had 4 haplotypes. All 4 haplotypes were found in the wingless aphid populations in May, with 7, 5, and 4 unique haplotypes in winged August, winged September, and wingless May populations, respectively (Fig. 3-B).

Fig. 1 Cytb and 16S gene haplotypes and variable sites of Aphis gossypii populations. The 46 variable sites were nt_6,nt_71, nt_119, nt_121, nt_123, nt_195, nt_198, nt_206, nt_212,nt_218, nt_245, nt_267, nt_269, nt_275, nt_276, nt_326,nt_329, nt_337, nt_339, nt_360, nt_370, nt_372, nt_404,nt_414, nt_433, nt_435, nt_449, nt_455, nt_470, nt_474,nt_488, nt_491, nt_515, nt_516, nt_518, nt_520, nt_523,nt_532, nt_538, nt_541, nt_542, nt_543, nt_544, nt_547, and nt_552.
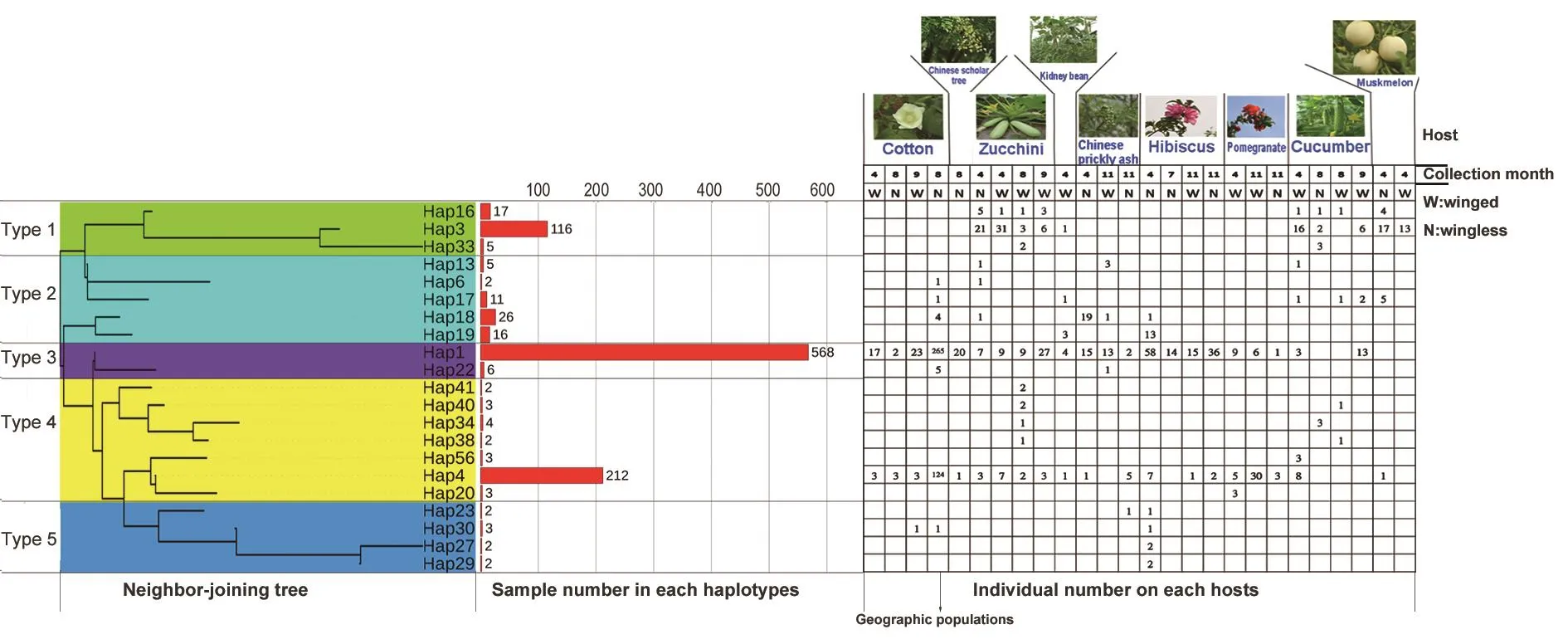
Fig. 2 Outline of 5 biotypes of Aphis gossypii in northern China. Five biotypes are colored according to the neighbor-joining tree,which was constructed using 21 haplotypes and associated with its host plant.
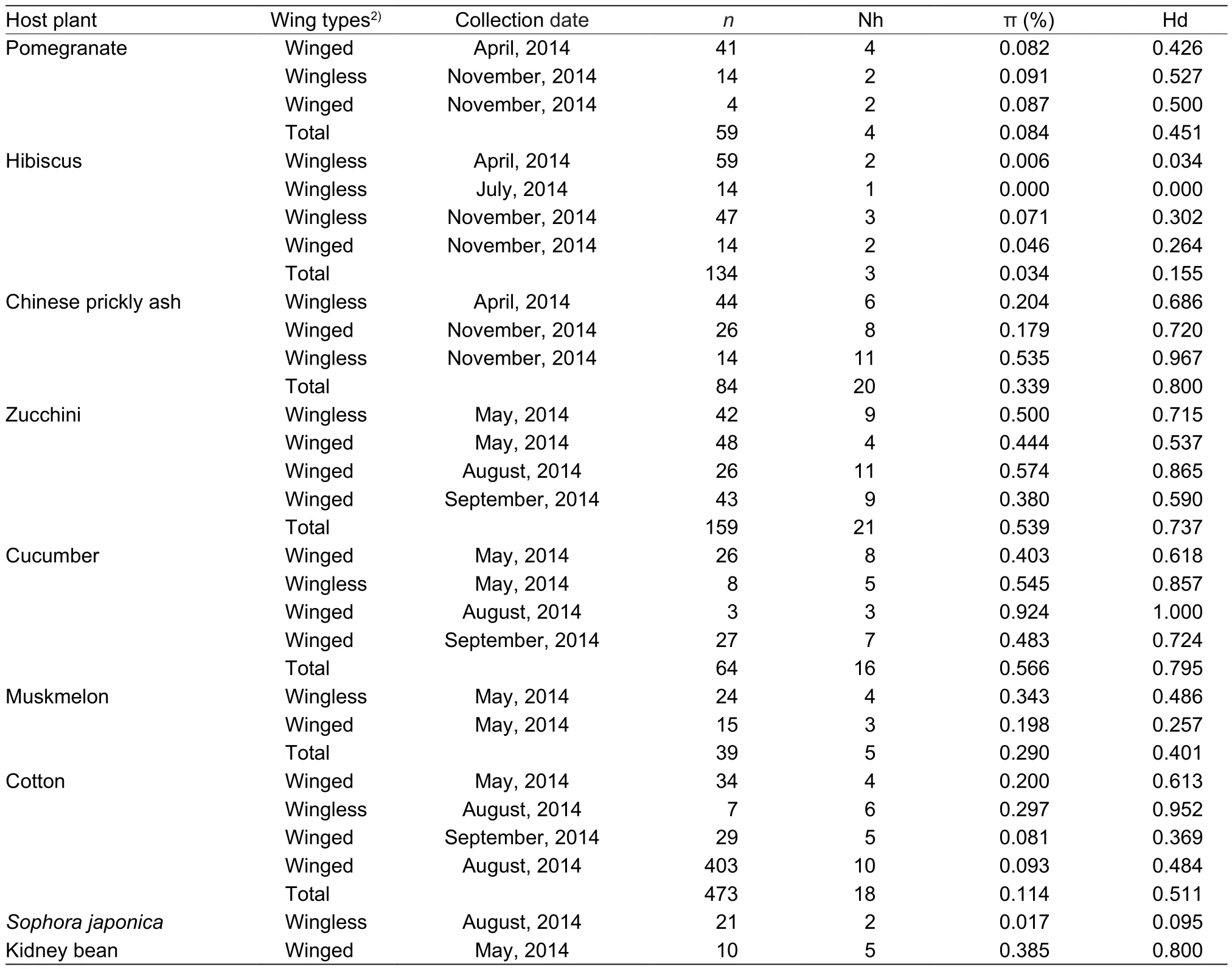
Table 1 Genetic diversity of 9 Aphis gossypii host-plant populations revealed by mitochondrial genes1)
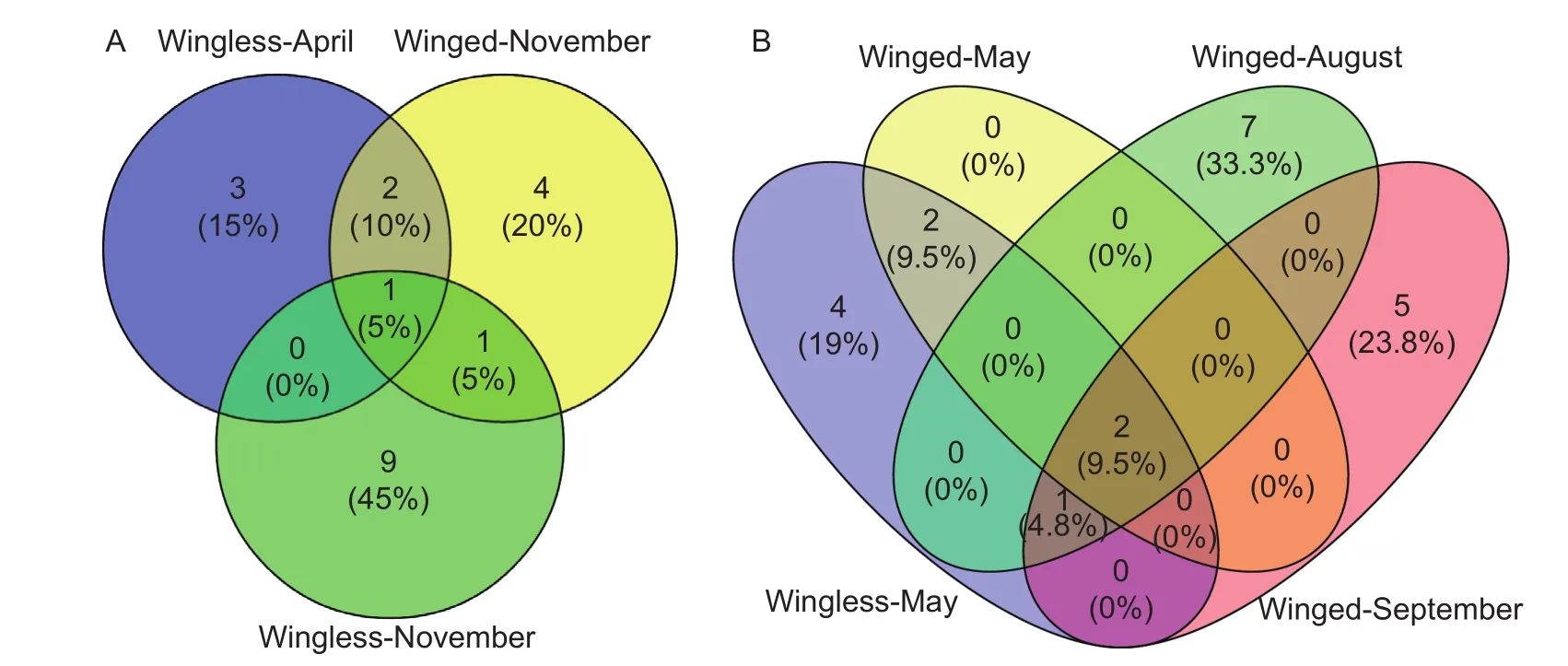
Fig. 3 Venn diagrams of the number of haplotype distribution across different Aphis gossypii populations. A, 3 aphid populations collected on Chinese prickly ash at 3 times. One common haplotypes was Hap1. B, 4 aphid populations collected on zucchini at 3 times. Four common haplotypes were Hap1, Hap3, Hap4 and Hap16. Wingless-April, wingless aphid populations in April;Wingless-November, wingless aphid populations in November; Winged-November, winged aphid populations in November;Winged-May, winged aphid populations in May; Winged-August, winged aphid populations in August; Wingless-May, wingless aphid populations in May; Wingless-August, wingless aphid populations in August.
3.3. ldentification of A. gossypii biotypes
Twenty-one haplotypes representing more than 2 individuals were chosen, and a NJ tree was constructed using MEGA Software. Associated with its host plant, 5 biotypes were identified. Type 1 contained 3 haplotypes, including Hap3,which was one of the top 3 most abundant haplotypes(Fig. 2). The host plants of Type 1 were zucchini, kidney bean, and muskmelon, the distinguishing feature was that it included no primary host. Type 2 contained 5 haplotypes,and was collected from 7 host plants. This biotype was rare on host plants except on Chinese prickly ash in April.Type 3 had the most individuals among the 5 biotypes, but only contained 2 haplotypes. Type 3 was collected from 8 host plants, and was abundant on each host plant, with the most abundance on cotton in August. The distinguishing feature of this biotype was that no individual was collected on muskmelon. Type 4 was the most abundant haplotype among the 5 biotypes, and was collected from all 9 host plants. Like Type 3, it was most abundant on cotton in August. Type 5 had the lowest abundance among the biotypes, being found in only 9 individuals collected on Chinese prickly ash and cotton. Individuals collected from S. japonica belonged to Type 3 and Type 4.
3.4. The biotypes of A. gossypii geographic populations
The haplotypes of 19 geographic populations collected from cottonfields were analyzed. The Nh ranged from 1 to 5. The average nucleotide diversity and Hd over all 19 populations were 0.0638% and 0.321, respectively. The populations collected from SDLL (Laoling, Shandong Province),HBLZ (Linzhang, Hebei Province), and SDSX (Shanxian,Shandong Province) only had 1 haplotype. The populations collected from SDJX (Jinxiang, Shandong Province) and HNYL (Yanling, Henan Province) had more abundance of haplotypes, with 5 and 4, respectively (Table 2).
All geographic populations and Anyang populations on cotton contained Type 3 individuals, 15 geographic populations and Anyang populations on cotton contained Type 4 individuals, 4 geographic populations contained Type 2 individuals, and 1 geographic population and Anyang populations on cotton contained Type 5 individuals (Fig. 4).
4. Discussion
A. gossypii may have undergone rapid diversification from the most recent common ancestor, and the population exhibited greater genotypic diversity as a result. Bioassay of laboratory host-transfer experiments was the original method used to confirm A. gossypii biotypes (Guldemond et al. 1994; Liu et al. 2008; Carletto et al. 2009b; Wang et al. 2016). Simple sequence repeat (SSR) markers were another popular method to assess A. gossypii diversity(Vanlerberghe-Masutti et al. 1999; Thomas et al. 2012; Luo et al. 2016). Mitochondrial gene sequence polymorphisms were also used to distinguish A. gossypii biotypes (Carletto et al. 2009a; Wang et al. 2016). Many studies have reported the existence of host-specific A. gossypii biotypes, but a lack of standardization has prevented comparisons of biotypes(Ebert and Cartwright 1997). In a previous study using SSRmarkers, we found that A. gossypii populations in northern China exhibited greater genotypic diversity, and the 31 populations in northern China could be classified into one of 3 biotypes according to host species (Luo et al. 2016). Unlike in other places, the microsatellite loci had high diversity in northern China, making it difficult to practically use. Based on complete mitochondrial sequences, a molecular marker with 5 single-nucleotide polymorphisms within Cytb and 16S genes was identified to distinguish the biotypes, and 2 biotypes of A. gossypii specializing on cotton and cucurbits in northern China were identified (Wang et al. 2016). Other studies also showed that the mitochondrial Cytb gene can be used to distinguish A. gossypii from related species in many places (Carletto et al. 2009a; Kim et al. 2011). In this study,we used single-nucleotide polymorphisms within Cytb and 16S genes beyond the 5 single-nucleotide polymorphisms used to distinguish A. gossypii biotypes, and these may be used in studying the life cycle of different host biotypes of A. gossypii in northern China.
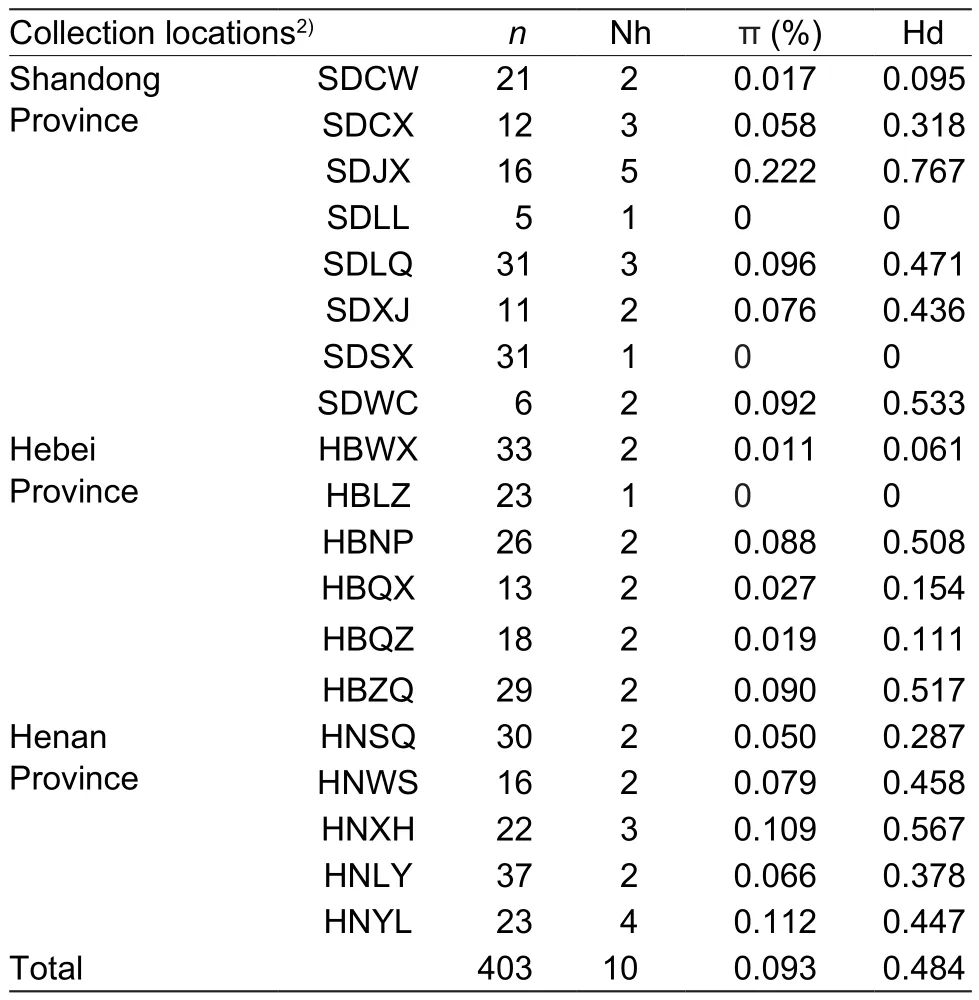
Table 2 Genetic diversity of 19 Aphis gossypii geographic populations in China revealed by mitochondrial genes1)
The cucurbit host-race A. gossypii has been well studied,and has been assigned to genetic clusters I, II, and III according to Vat resistance of Cucumis melo (Boissot et al.2016; Thomas et al. 2016). Type 1 A. gossypii exactly corresponded to the cucurbit host-race. Type 1 contained 3 haplotypes in northern China, and was only found on zucchini, kidney bean, cucumber and muskmelon. No primary hosts were found in this study. Zucchini and kidney bean have been shown in previous studies to serve as hosts without consideration of the host-race of A. gossypii(Wu et al. 2013). Results indicated that cucurbit host-race can be anholocyclic or holocylic lineages and originating from wild plants that had never been previously sampled(Thomas et al. 2012). The haplotypes that contained more than 2 individuals were chosen to construct a NJ tree and to identify A. gossypii biotypes, as less abundant haplotypes could be derived from nucleotide incorporation mistakes that occurred during the PCR or sequencing procedure (Biles and Connolly 2004).
Unlike the cucurbit host-race that was not found on cotton, 3 out of 4 biotypes considered in the cotton hostrace of A. gossypii were on cucurbits. In melon-growing areas offrance, Bayesian Structure results showed that a cluster contained some multilocus genotypes (MLGs) that specialized on Cucurbitaceae and some specialized on Malvaceae (Thomas et al. 2012).
In this study, zucchini was planted near cotton and Cucurbitaceaefields, and the cotton host-race may have transferred from cotton to Cucurbitaceae with zucchini as a bridge plant (Wu et al. 2013). A large scalefield experiment in a Cucurbitaceae plantfield is needed in order to confirm this hypothesis.
A. gossypii can be holocyclic in northern China. The other 4 biotypes in the cotton host-race have this lifecycle because allof them have both primary and summer hosts.Those primary hosts are not only hibiscus, pomegranate,and Chinese prickly ash, but also S. japonica, which was reported for the first time in China to our knowledge. Using classification with the mitochondrial gene, pomegranate was the host of the cotton host-race, and when using SSR, the population on pomegranate was classified with populations collected on cotton (Luo et al. 2016). The Chinese prickly ash (Rutaceae) is one of the most ancient cultivated economic trees in Chinese mountainous regions, and is of major agricultural importance as sources of spices and traditional medicine (Feng et al. 2015). All 4 biotypes of A. gossypii can be found on Chinese prickly ash, with Type 5 being on cotton and Chinese prickly ash. The highest haplotype diversity value was observed in populations on Chinese prickly ash, but populations from globally distributed hibiscus had lower diversity. Since Chinese prickly ash originated from China and has a narrow range, the A. gossypii immigrant colony may host on this plant, leading to a greater genetic diversity.
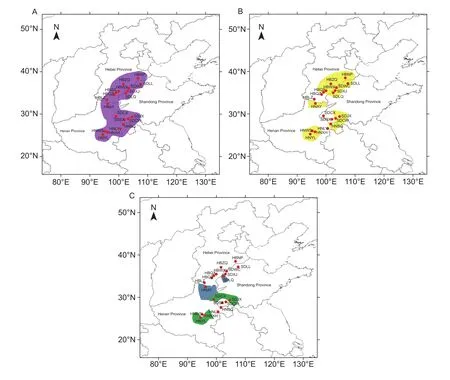
Fig. 4 Distribution of Aphis gossypii biotypes over 20 A. gossypii geographic populations in 3 provinces in China. A, Type 3.B, Type 4. C, Type 2 (green) and Type 5 (blue). See Table 2 for detailed information of all the abbreviations of geographic populations.
Nineteen geographic populations were collected in the North China Plain beside Anyang. Those places are traditional cotton planting areas, and the cotton in collectionfields are far from other A. gossypii hosts such as zucchini and cucumber. Allof the individuals belonged to the 4 biotypes that were classified in the cotton host-race (Luo et al. 2016). Generally, Type 3 was the main biotype in cottonfields in northern China.
5. Conclusion
In this study, we used single-nucleotide polymorphisms within the Cytb and 16S genes beyond the 5 singlenucleotide polymorphisms previously used to distinguish A. gossypii biotypes in northern China. The life cycles of different host biotypes of A. gossypii in northern China were discussed. Type 3 was the main biotype found in northern China cottonfields.
Acknowledgements
This research was supported by the National Natural Science Foundation of China (31572015) and the National Special Transgenic Project of China (2016ZX08012-004).
杂志排行

Journal of Integrative Agriculture的其它文章
- ldentification and characterization of Pichia membranifaciens Hmp-1 isolated from spoilage blackberry wine
- Implications of step-chilling on meat color investigated using proteome analysis of the sarcoplasmic protein fraction of beef longissimus lumborum muscle
- Spatial-temporal evolution of vegetation evapotranspiration in Hebei Province, China
- Design of a spatial sampling scheme considering the spatialautocorrelation of crop acreage included in the sampling units
- Comparison of forage yield, silage fermentative quality, anthocyanin stability, antioxidant activity, and in vitro rumen fermentation of anthocyanin-rich purple corn (Zea mays L.) stover and sticky corn stover
- Synonymous codon usage pattern in model legume Medicago truncatula