Effect of pre-culture on virus elimination from in vitro apple by thermotherapy coupled with shoot tip culture
2018-08-06HUGuojunDONGYafengZHANGZunpingFANXudongRENFangLIZhengnan
HU Guo-jun, DONG Ya-feng, ZHANG Zun-ping, FAN Xu-dong, REN Fang, LI Zheng-nan
Research Institute of Pomology, Chinese Academy of Agricultural Sciences, Xingcheng 125100, P.R.China
Abstract We evaluated the role of pre-culture on survival rate of in vitro apple plants treated by thermotherapy. Two apple cultivars,Malus×domestica cv. Pink Lady and Huafu, were used in the experiment and both have widely grown in China and infected with Apple chlorotic leaf spot virus (ACLSV) and Apple stem grooving virus (ASGV). Results in growth and virus titer of apple plants did not exhibit clear trends duringfive different periods of pre-culture. Whilst, pre-culture increased the survival rate of the two cultivars during thermotherapy. The survival rate of plants pre-cultured for 13 d (P-13d) was 14 and 51% higher than that of P-1d plants for Pink Lady and Huafu, respectively. Moreover, pre-culture positively in fluenced regeneration of Huafu plants. The average survival rate of plants regenerated from P-1d and P-4d was 20% lower than that regenerated from P-7d, P-10d, and P-13d. The efficiency of virus eradication was determined by reverse-transcription PCR with two primer pairs for each virus, and the detection results showed that pre-culture scarcely affected apple virus elimination.Despite the fact that the two viruses were hardly detected at 5 d of thermotherapy, no virus-free plants were found in the two cultivars of regenerated apple plantlets after 30-d treatment.
Keywords: apple, in vitro culture, pre-culture, thermotherapy, virus elimination
1. Introduction
The apple (Malus×domestica) belongs to the family Rosaceae which is one of the most widely cultivated temperate fruit trees. China is one of the main apple-producing countries,producing almost half of the world’s apples with a planting area of 2.3 million ha and a production of 41 million tons offruit in 2014 (Hu et al. 2016). Genetic resources are required to breed new apple cultivars in both traditionaland genetic engineering breeding programs. An alternative breeding method is vegetative propagation; however, viruses are particularly problematic in vegetatively propagated crops because they are transmitted from generation to generation.Apple chlorotic leaf spot virus (ACLSV, genus Trichovirus,family Beta fl exiviridae), Apple stem pitting virus (ASPV,genus Foveavirus, family Beta fl exiviridae) and Apple stem grooving virus (ASGV, genus Capillovirus, family Beta fl exiviridae) are common viruses and infected plants are generally asymptomatic (Adams et al. 2004). They significantly reduce fruit yield and quality, and commonly occur as mixed infections, causing top-working disease of apple on susceptible rootstocks (Yanase 1983; Desvignes and Boyé 1988). All these viruses are naturally disseminated via infected propagation materials (Yanase et al. 1989; Pasquini et al. 1998; Hirata et al. 2003). Apple is vegetatively propagated by grafting, thus resulting in virus transmission from generation to generation.
Thermotherapy is an efficient technique to eliminate viruses from infected apple plants (Paprstein et al. 2008;Hu et al. 2015a, b). A higher temperature over an extended time period can reduce the movement of virus particles into the apical meristem by inhibiting viral RNA synthesis (Valero et al. 2003; Wang et al. 2008). Even if the viruses are not eliminated, higher temperatures can sufficiently slow down their replication and infection rates to allow new shoots to grow (Wang et al. 2008). However, studies have reported that prolonged heat treatments would severely negatively affected the growth and survivalof apple plants (Paprstein et al. 2008; Hu et al. 2015b).
In previous researches, Hu et al. (2015a) found that the duration of pre-culture may have effects on the survivalof high temperature-treated plants. In this study, we investigated the effects of pre-culture duration on the survival rate and viral elimination in in vitro samples of two apple cultivars that were also exposed to a thermotherapy treatment. We hypothesized that pre-culture and thermotherapy acting to gether would increase the viral elimination and plant survival. We compared virus concentrations in in vitro apple plants that were pre-cultured at one offive different time periods to determine the percentage of viral elimination.Plant survival was determined by calculating the percentage of initial shoots after thermotherapy.
2. Materials and methods
2.1. Plant materials
Two apple cultivars that are widely grown in China, in vitro Malus×domestica cv. Pink Lady and Huafu, were used as the plant materials in the experiment. Shoots of the two cultivars were collected in spring from the Research Institute of Pomology, Chinese Academy of Agricultural Sciences.The presence of ACLSV and ASGV was tested by reverse transcription PCR (RT-PCR).
2.2. In vitro cultures
In vitro plants were cultured on modified Murashige and Skoog(MS) medium containing 1.0 mg L-16-benzylaminopurine,0.2 mg L-1indole-3-butytric acid, 30 g L-1sucrose, and 5.6 g L-1agar. The pH was adjusted to 5.8 with 1 mol L-1NaOH or HCl before autoclaving at 121°C for 15 min. All cultures were kept in a standard growth room at (24±1)°C under a 16 h light/8 h dark photoperiod with 2 000 lx light intensity.Viable cultures were transferred to fresh MS medium at 30-40 d intervals, and the presence of ACLSV and ASGV in the cultures was reassessed by RT-PCR.
2.3. Isolation of RNA and RT-PCR
Total RNA was extracted from apple samples as described by Hu et al. (2017). First-strand cDNA synthesis was conducted in a 20-μL reaction containing approximately 1 μg RNA, 5× M-MLV RT buffer, 0.5 mmol L-1dNTP mix(Roche Diagnostics, Mannheim, Germany), 5 μmol L-1random hexamer primers (TaKaRa, Dalian, China), and 5 U μL-1M-MLV reverse transcriptase (Promega, Madison,WI, USA). The mixtures were incubated at 37°C for 10 min and then at 42°C for 1 h. The PCR was carried out in a 25-μL reaction volume containing 10× PCR buffer with Mg2+,10 mmol L-1dNTP mix, 0.5 μL each primer (final concentration 1 μmol L-1), 0.2 μL rTaq DNA polymerase (U μL-1) (TaKaRa),2 μL template cDNA, and sterile water. Each apple virus was detected using two primer pairs (Table 1). The cycling profile comprised a 3-min incubation at 94°C, followed by 35 cycles of 94°C for 30 s (denaturation), 50-55°C for 30 s(annealing), and 72°C for 30-50 s (extension), with afinal extension step at 72°C for 10 min. Amplicons were separated by electrophoresis on 1.2% (w/v) agarose gels,stained with ethidium bromide, and then visualized under ultraviolet light.
2.4. Quantitative real-time PCR (qRT-PCR)
Total RNA samples were treated with the RT reagent kit with gDNA Eraser (TaKaRa) to remove genomic DNA and synthesize the first strand cDNA according to the manufacturer’s instructions. In a PCR tube, 2.0 μL 5×gDNA Eraser buffer, 1.0 L gDNA Eraser, and 3.0 μg total RNA were mixed and then RNase-free dH2O was added up to 10 μL, and the solution was then incubated at 42°C for 2 min. Additional reagents of 4.0 μL 5× PrimeScript buffer,1.0 μL PrimeScript RT Enzyme Mix I, and 1.0 μL RT Primer Mix was then added to the previous tube, then RNase-free dH2O was added up to 20 μL, and the solution was then incubated at 37°C for 15 min followed by 85°C for 5 s and stored at −20°C for later use.
The titres of ACLSV and ASGV during pre-culture and thermotherapy was quantified using SYBR®Premix Ex Taq™II (TaKaRa) according to the manufacturer’s instructions on the CFX Connect Real-Time System (Bio-Rad, Hercules,CA). The 20 μL reaction mixtures were incubated at 95°C for 2 min for the initial denaturation stage, followed by 40 cycles at 95°C for 10 s, 58°C for 15 s and 72°C for 20 s. The ΔCtmethod was used to calculate the relative concentration of ACLSV and ASGV during treatment. Each sample was examined in triplicate. The primers used in this study were synthesized by Sangon Biotech (Shanghai, China). A primer pair that allowed for the specific amplification of mRNA corresponding to the mitochondrial nad 5 (Ma et al. 2008)gene was used as an internal control (Table 1).
2.5. Pre-culture and thermotherapy
Shoots approximately 6.0 mm long were excised from in vitro cultures and were placed into culture bottles containing fresh MS medium. The plantlets were pre-cultured in a growth room for 1, 4, 7, 10, and 13 d, respectively and plant materials were collected after each pre-culture period.
After pre-culture, the culture bottles were moved into a heated chamber. The temperature was raised at 3°C d-1,then the cultivated temperatures were as follows: 27, 30,33, and 37°C, and the thermotherapy periods were recorded when the temperature reached 37°C. The duration of all thermotherapy treatments was 30 d. Plant materials were collected at different culture temperatures of 27, 30, 33, and 37°C, and collected at 37°C for 0, 1, 5, 10, 15, 20, 25, and 30 d of thermotherapy, respectively.
The growth and heights of the in vitro apple plants were observed periodically to evaluate the effects of the different treatments. In total, 45 shoots were used for each treatment.Meristem tips approximately 1.0 mm long were dissected from apical shoots after treatments and cultured on the same basic MS medium as that described above. Cultures from the same source kept in a standard growth room were used as controls. After three cycles of sub-culture, RT-PCR was conducted to detect viruses, and the efficiency of virus elimination was calculated as follows: Number of virus-free plants/Number of initial shoots.
3. Results
3.1. Effect of pre-culture on growth of in vitro apple plantlets
All in vitro Pink Lady and Huafu plants survived and grewedhealthy during pre-culture (Table 2). The heights of plants for the two cultivars showed hardly any difference among the five pre-culture periods, but the leaves of plants pre-cultured for 13 d (P-13d) was slightly bigger than that of P-1d (Fig. 1).Moreover, as the period of pre-culture time extended, the increase in size of basal callus was more obvious in the cultivar Pink Lady. After 13 d of pre-culture, there were no new shoots observed in both cultivars (Fig. 1).
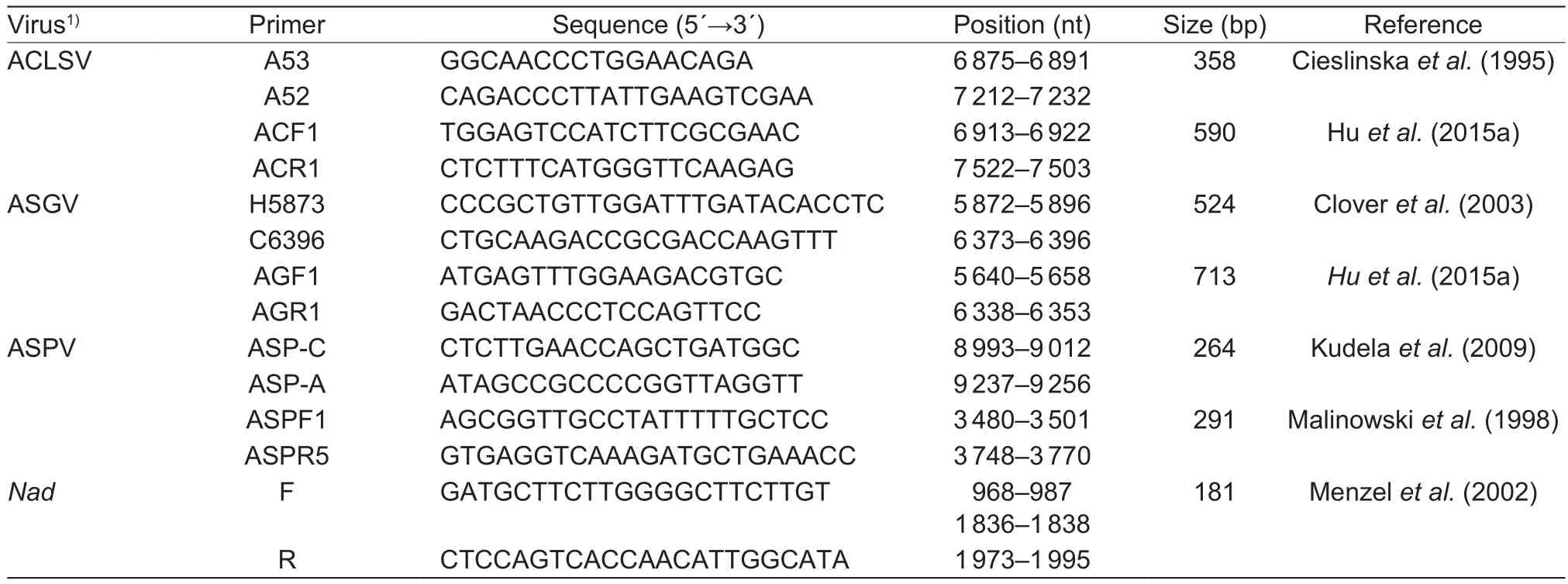
Table 1 Oligonucleotide primers used in reverse transcription PCR (RT-PCR) assays to detect apple viruses

Table 2 Survival rate after pre-culture, thermotherapy and shoot tip culture
3.2. Effect of pre-culture on apple virus titer
The presence of ACLSV and ASGV in Pink Lady and Huafu plants was detected using a standard RT-PCR after 1, 4, 7,10 and 13 d of pre-culture. Non-specific bands were rarely observed, and no amplicons were generated for the negative control samples. We observed that targets of ACLSV and ASGV showed little difference among the five periods of pre-culture (Fig. 2). The relative quantities of ACLSV and ASGV in the two cultivars were verified by qRT-PCR, and the results of the titers of the two viruses lacked any increasing or decreasing trends with duration of pre-culture, which indicated that the regularity of virus-titer change in different periods was unclear (Fig. 3).
3.3. Effect of thermotherapy on growth of pre-culture apple plantlets

Fig. 1 Growth of in vitro apple plants after pre-culture for 1, 4, 7, 10 and 13 d (P-1d, P-4d, P-7d, P-10d, and P-13d), respectively.A, in vitro Pink Lady plants. B, in vitro Huafu plants.
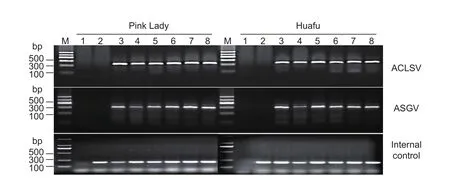
Fig. 2 Agarose gel electrophoresis of reverse transcription PCR (RT-PCR) products of Apple chlorotic leaf spot virus (ACLSV)and Apple stem grooving virus (ASGV) in the five different periods of in vitro pre-culture and an internal control. M, DNA marker II(Tiangen, Beijing, China); lane 1, no template control; lane 2, negative control; lane 3, no positive control; lanes 4-8, pre-culture for 1, 3, 7, 10 and 13 d, respectively. The primer sets for ACLSV and ASGV detection were A53/A52 and H5873/C6396, respectively.
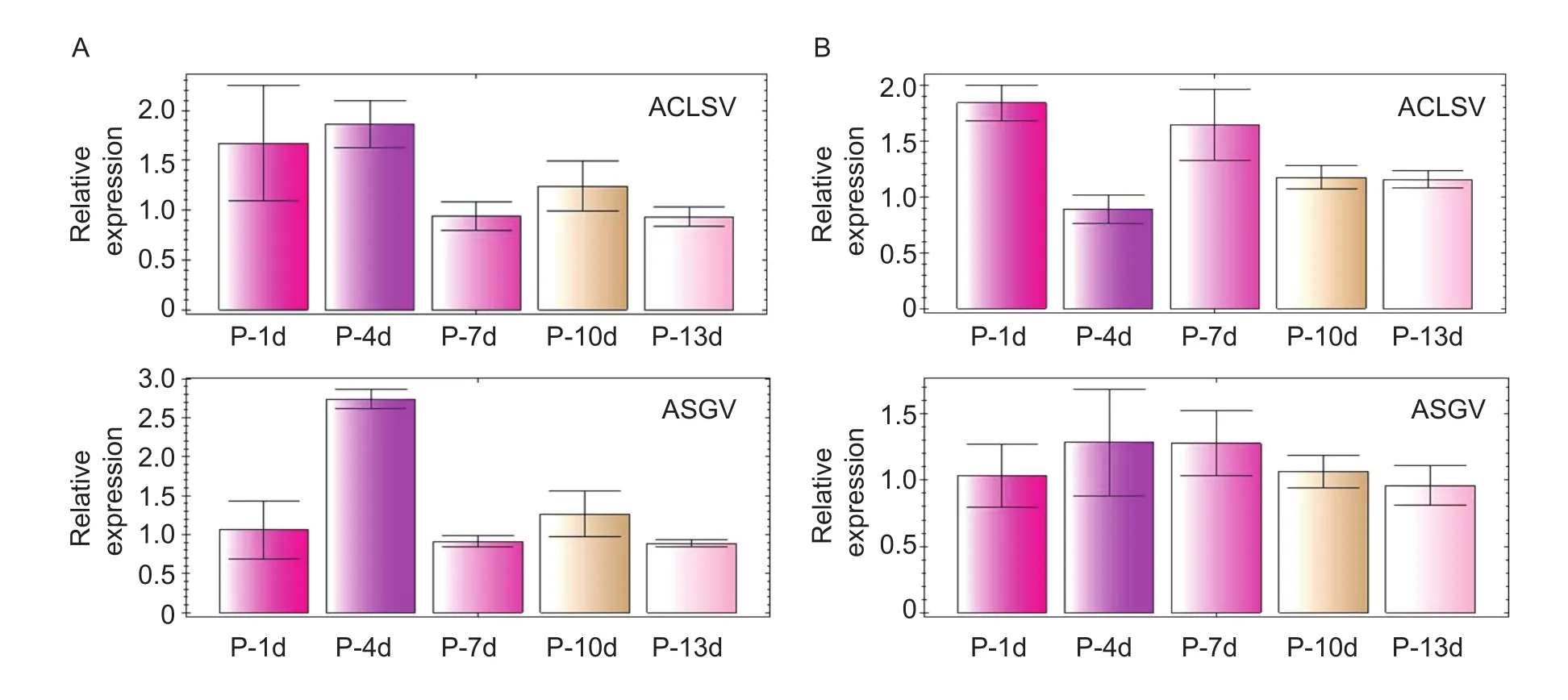
Fig. 3 Quantitative reverse transcription PCR (RT-PCR) was performed to quantify the relative expression levels of Apple chlorotic leaf spot virus (ACLSV) and Apple stem grooving virus (ASGV) in in vitro Pink Lady (A) and Huafu (B) plants after pre-cultured for 1, 4, 7, 10 and 13 d (P-1d, P-4d, P-7d, P-10d, and P-13d), respectively. Error bars represent the standard deviations among triplicate experimental reactions.
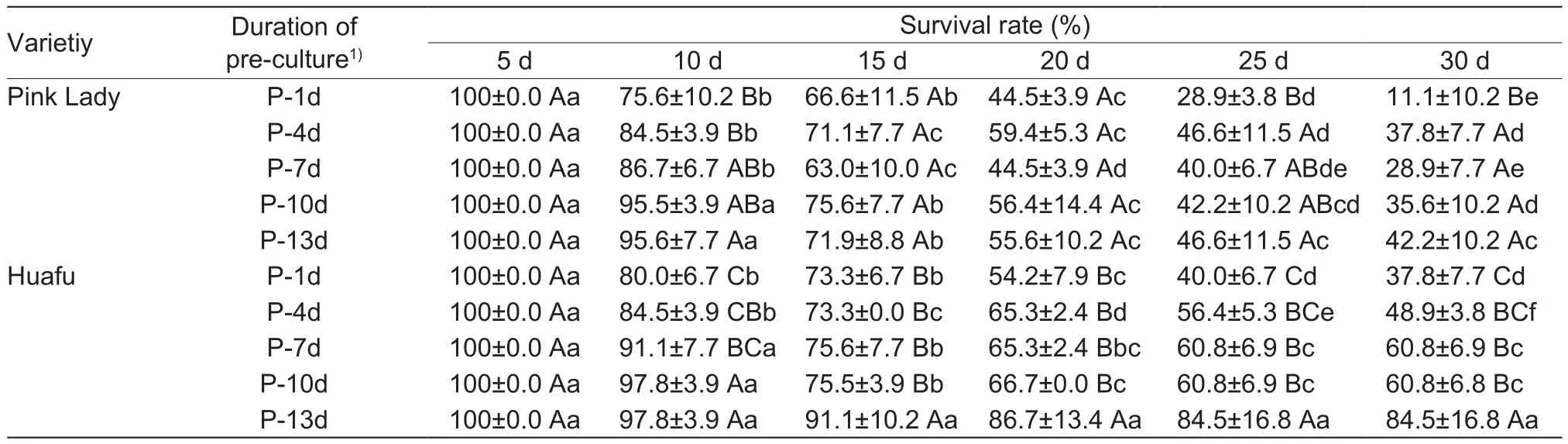
Table 3 Survival rate of apple plants during thermotherapy
The survival rate of in vitro Pink Lady and Huafu plants decreased as the duration of thermotherapy increased, but the pre-culture treatment generally had positive effects on the growth of the two cultivars (Table 3). For Pink Lady,the survival rate of P-1d plants was lower than that of P-4d,P-7d, P-10d, and P-13d plants when thermotherapy lasted 10, 25 and 30 d. Onlyfive plants survived after 30 d of thermotherapy (Table 3). The effect of high temperature on plants among the five pre-culture periods was not obvious during 20 and 25 d of thermotherapy. At the end of thermotherapy, the leaves of P-1d, P-4d and P-7d plants turned yellow, brown or black and dessicated; the tips of most plants became brown or black. Moreover,growth of plants in these three treatments was nearly similar to that of the control plants, and the growth speed of P-10d and P-13d plants was higher than that of the former three treatments (Fig. 4-A). For Huafu, the survival rate of P-13d plants was higher than that of P-4d,P-7d and P-10d plants during the longest thermotherapy treatment (30 d), and there was no significant difference among different durations of the thermotherapy. The percentage of surviving plants was up to 82.2% at the end of thermotherapy (30 d) (Table 3). When the treatment lasted 30 d, the leaves of P-1d and P-4d plants turned yellow and brown and became desiccated. No growth was observed in these two treatments when compared with the control plants and the growth of P-7d and P-10d plants was higher than that of the former two treatments. The P-13d plants showed almost the same amount of growth with that of control (Fig. 4-B). Shoot proliferation was also strongly inhibited by higher temperatures. Almost no axillary shoots regenerated for the two cultivars in the 37°C treatments.
3.4. Effect of pre-culture on survival rate of shoot regeneration
In vitro Pink Lady and Huafu plants were regenerated from treated shoot tips (1.0 mm) and the survival rate was calculated (Table 2). The effect of pre-culture duration on regeneration of Pink Lady was not obvious, and no clear trend was observed for this cultivar. Whereas, pre-culture clearly affected regeneration of Huafu plants. The average survival rate of plants regenerated from P-1d and P-4d was 20% lower than that regenerated from P-7d, P-10d and P-13d.
3.5. Effect of thermotherapy on ACLSV and ASGV titers
The presence of ACLSV and ASGV in the high temperaturetreated Huafu plants was detected using a standard RTPCR when the temperature raised to 27, 30, 33 and 37°C,respectively, and during 37°C for 0, 1, 5, 10, 15, 20, 25, and 30 d, respectively. There was no band for the two viruses after 5 d at 37°C. We speculated that the viral titer would decrease as early as the start of the treatment (Fig. 5).
3.6. Efficiency of virus eradication by thermotherapy combined with shoot tip culture
Regenerated plants sub-cultured for three cycles were analyzed by RT-PCR to confirm the elimination of viruses,using two primer pairs for each virus. Plants were judged as virus-free when there were no target products amplified by RT-PCR with four primer pairs. Virus elimination efficiencies differed in the two apple cultivars (Table 4). The average elimination rates of ACLSV and ASGV in regenerated Pink Lady plants was 21.7 and 8.7%, respectively. Only two regenerated plants were ASGV-free. Almost no ACLSV was eliminated from regenerated Huafu plants, and the average eradication rate of ASGV was 24.2%. The pre-culture scarcely showed any effect on the apple virus elimination rate. The detection results showed that no virus-free plants were found in the two regenerated apple cultivars.

Fig. 4 Growth of in vitro apple plants after thermotherapy for 30 d. A, in vitro Pink Lady plants. B, in vitro Huafu plants. CK,in vitro apple plants were cultured in a standard growth room; P-1d, P-4d, P-7d, P-10d, and P-13d, pre-cultured for 1, 4, 7, 10,and 13 d, respectively.

Fig. 5 Agarose gel electrophoresis of reverse transcription PCR (RT-PCR) products of ACLSV (Apple chlorotic leaf spot virus)and ASGV (Apple stem grooving virus) infive different periods of thermotherapy and a internal control in Huafu plants. M, DNA Marker II (Tiangen, Beijing, China); lane 1, no template control; lane 2, negative control; lane 3, no positive control; lanes 4-6,temperature at 27, 30, and 33°C, respectively; lanes 7-13, pre-cultured for 1, 5, 10, 15, 20, 25 and 30 d (P-1d, P-3d, P-7d, P-10d,and P-13d) at 37°C, respectively. The primer sets for ACLSV and ASGV detection were A53/A52 and H5873/C6396, respectively.
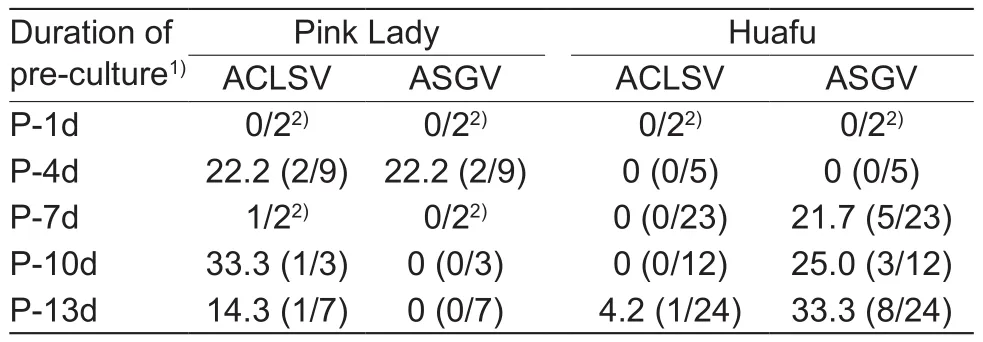
Table 4 Efficiencies of thermotherapy to generate Apple chlorotic leaf spot virus (ACLSV)- or Apple stem grooving virus(ASGV)-free apple plants
4. Discussion
Thermotherapy is a routine method for virus eradication,and this method has been widely used to eliminate viruses from fruit trees and other vegetative plants (Valero et al.2003; Paprstein et al. 2008; Tan et al. 2010). Woody plants,like apple, are sensitive to prolonged exposure to extreme high temperatures (Paprstein et al. 2008; Tan et al. 2010;Hu et al. 2012). High temperatures significantly affected shoot survival, and the survival rate directly associated with the success of thermotherapy. In this research, we tried to investigate the potential in fluence of various durations of pre-culture to increase the survival rate of apple plantlets.The success of virus elimination is also related to the concentration of each particular virus in the plant material. It was unclear whether the titers of virus would change during pre-culture, so the relative quantitative concentration of ACLSV and ASGV in the Pink Lady and Huafu plants were analyzed. The results showed no obvious trend of the two virus’s titers in the two cultivars during pre-culture. Thus, we speculated that pre-culture did not affect the virus titers, and the differences among the five periods were due to individual differences between plantlets. It showed that the longer time pre-culture could increase the growth of plants, which were helpful for the plants during high temperature treatment.
The temperature range is an important factor affecting shoot survival. Most often, the temperature of thermotherapy is measured in a growth chamber. Some authors have shown that the air temperature measured in the growth chamber was 1 or 2°C (Deogratias et al. 1989; Stein et al. 1991) lower than that inside the test tubes within the chamber. When choosing a temperature, there must be a balance between relatively low shoot mortality at higher temperatures and the high percentage of virus-free shoots (Dziedzic 2008).Previous research demonstrated that the survival rates of shoots from apple plants treated at 34 and 36°C was 84.6 and 93.3%, respectively, but almost no shoots survived in the 38°C treatment (Hu et al. 2015a). We chose 37°C to eliminate the apple viruses, and the results indicated that this temperature caused high death rates, thus, the method of pre-culture was applied to determine whether it can mitigate the often fatal effect of high temperatures. The survival rate of P-13d plants was 14 and 51% higher than that of P-1d plants for Pink Lady and Huafu, respectively.However, although the in vitro apple plants were cultured in the same conditions, inherent variability among individuals within each cultivar may be responsible for the differences we observed, for example, in the P-4d plants of Pink Lady and P-10d plants of Huafu.
The duration of thermotherapy treatment can affect virus elimination efficiency (Tan et al. 2010; Hu et al. 2012). Dong et al. (2002) found that the elimination rate of an apple virus treated with a high temperature of 37°C for 36 d was clearly higher than that of 27 d. Cheng et al. (2003) had reported that when a 38°C temperature lasted for 25 d, the elimination rates of ACLSV and ASGV were 17% higher than that lasted for 20 d. In the present study, the duration of the high temperature treatment was 30 d long, and surviving plants(11.1-82.2%) were found in all pre-culture treatments at the end of treatment. However, the effects of temperature and duration of exposure may be obscured by the intrinsic heat tolerance of the host plant because the levelof tolerance depends on species and variety (Hu et al. 2015b). Pink Lady plants were more sensitive to high temperature than that of Huafu.
The results showed that pre-culture had little effect on apple virus elimination. Unfortunately, none of the regenerated apple plants were virus-free. In fact, the titers of ACLSV and ASGV during thermotherapy were detected,and no virus was found when thermotherapy lasted for 5 d.The underlying mechanism of thermotherapy’s effect on reducing viral infection has not been completely explained yet. Since Kunkel (1935) established that exposure of virus-infected trees in pots to temperatures ranging between 34.4 and 36.3°C was effective in treating peach tree of either peach yellows or peach rosette, heat treatment for the inactivation of different viruses in plants has become an established practice. One explanatory hypothesis is that heat stress, especially at sublethal temperatures, inhibits virus replication in newly growing plant tissues and thus allows a plant to recover from viral infections (Nyland and Goheen 1969; Mink et al. 1998). Szittya et al. (2003) and Qu et al. (2005) reported that plant-virus interactions are strongly affected by temperature, and high temperatures are frequently associated with low virus content in infected plants. Wang et al. (2008) revealed a drastic decrease in the amounts of Raspberry bushy dwarf virus RNA in shoot tips and leaves following 5 d of thermotherapy; moreover,the amounts of viral RNA were barely detectable following 8 d of thermotherapy. The plants likely did not have time to grow significantly in 8 d and the tissues sampled at different time points were essentially the same developmentally.They speculated that it may be explained by enhanced RNA silencing, the cellular mechanism devoted to targeting and degrading viral RNA in plants, at elevated temperatures.Our results supported the same conclusion.
Dong et al. (2002) reported 65% virus eradication from virus-infected apple plants using thermotherapy for 27 d.In the present study, none of the virus-free apple plantlets regenerated after high temperature treatment. The result also differed from our previously reported results (Hu et al.2015a), in which 43.7 and 52.4% of the regenerated plants from (34±0.5)°C and (36±0.5)°C treatments were virusfree. The reasons for this difference in virus elimination efficiency are not known, although factors such as the use of shoot tips from in vitro plants as wellas the different apple variety, and, perhaps, virus strain and concentration may have played a role in determining the differences in results.Moreover, the detection method also may have caused this difference (Bayati et al. 2011). In this study, RT-PCR has been used for virus detection, and to ensure accuracy, two primer pairs for each apple virus were used to increase the specificity and sensitivity of detection, while Dong et al.(2002) used enzyme-linked immuno-sorbent assay (ELISA)and indicator plants as a detection method.
5. Conclusion
This experimental modelallowed us to evaluate the modifications of thermotherapy. It proved that some improvement was needed to increase the virus elimination rate of different elimination methods. Furthermore, this study also lends further insight into the underlying mechanisms of thermotherapy.
Acknowledgements
This work was supported by the Agricultural Science and Technology Innovation Program of the Chinese Academy of Agricultural Sciences (CAAS-ASTIP).
杂志排行

Journal of Integrative Agriculture的其它文章
- ldentification and characterization of Pichia membranifaciens Hmp-1 isolated from spoilage blackberry wine
- Implications of step-chilling on meat color investigated using proteome analysis of the sarcoplasmic protein fraction of beef longissimus lumborum muscle
- Spatial-temporal evolution of vegetation evapotranspiration in Hebei Province, China
- Design of a spatial sampling scheme considering the spatialautocorrelation of crop acreage included in the sampling units
- Comparison of forage yield, silage fermentative quality, anthocyanin stability, antioxidant activity, and in vitro rumen fermentation of anthocyanin-rich purple corn (Zea mays L.) stover and sticky corn stover
- Synonymous codon usage pattern in model legume Medicago truncatula