Clostridium clariflavum GH10木聚糖酶的克隆表达、酶学性质及位点功能分析
2018-07-18王华广刘雨露胡方觊唐蕾
王华广,刘雨露,胡方觊,唐蕾,2*
1(江南大学 工业生物技术教育部重点实验室,江苏 无锡,214122) 2(江南大学 生物工程学院,江苏 无锡,214122)
木质纤维素是丰富的可再生生物质资源,占地球所有可再生有机碳的1/3[1]。目前这部分潜在资源并未被有效开发利用,造成了生物质资源的极大浪费[2]。木聚糖是半纤维素的主要成分,如何降解木聚糖是利用半纤维素生物质资源的关键。木聚糖酶可以水解木聚糖中的糖苷键,把木聚糖降解成木糖或者低聚木糖。大多数木聚糖为杂多糖侧链上含有不同的基团,如阿拉伯糖、乙酰基、阿魏酸残基、4-O甲基葡萄糖醛酸基等[3]。因此完全水解木聚糖需要多种酶的共同作用,其中起重要作用的是β-1,4内切木聚糖酶。生物酶法催化有效率高和环境友好的优点,在工业应用中正在取代化学催化法。木聚糖酶主要应用在纺织、食品、饲料、造纸、木质纤维素生物质转化等工业方面[4]。由于不同应用环境对酶有不同的要求,因此开发出适用于不同用途的木聚糖酶非常必要,目前采取的手段包括新酶的挖掘和酶的分子改造[5-7]。
根据蛋白的氨基酸序列、结构特征、底物的特异性、作用机制等理化性质的不同,木聚糖酶可分为6个糖苷水解酶家族:GH5、GH8、GH10、GH11、GH30、GH43,大多数木聚糖酶属于GH10和GH11[8-9]。GH10家族木聚糖酶具有较大的分子质量(>30 kDa)、低等点电、广泛的底物特异性、典型的(β/α)8桶折叠结构等特点[5,10],是近几年来研究与应用的热点之一,如GH10家族新酶的寻找、非理性改造、理性设计、结构解析、作用机制、末端氨基酸的作用等方面[11-16]。
Clostridiumclariflavum是一种新发现的嗜热厌氧木质纤维素降解菌,基因组学和蛋白质组学表明,C.clariflavum菌拥有丰富的木质纤维素降解酶系种类,在木质纤维素的降解方面有广泛的应用潜力[17]。目前关于此菌来源的重组酶在国内外的研究鲜有报道。本课题组之前首次在大肠杆菌中克隆表达了C.clariflavum的一个双催化域木聚糖酶基因Clocl-2441(结构域为GH11-CBM6-DockerinI-GHl0),并初步研究了其酶学性质[18]。本研究将Clocl-2441中的单个GH10结构域构建到pET28a(+)载体上,研究其酶学性质,并通过分子改造发现209位点的组氨酸是酶活性的重要氨基酸,研究结果为具有双催化域的木聚糖酶的分子改造和应用提供了依据。
1 材料与方法
1.1 材料与试剂
1.1.1 菌种与质粒
大肠杆菌(Escherichiacoli) JMl09、BL21(DE3)宿主菌株以及pET28a(+)、pET28a(+)-2441质粒均为实验室所保藏。
1.1.2 酶与试剂
限制性内切酶(EcoR I,Hind III)、T4DNA连接酶、高保真DNA聚合酶、质粒提取试剂盒、蛋白定量试剂盒,均购自TaKaRa(大连)。山毛榉木聚糖购自爱尔兰Megazyme公司。
1.2 方法
1.2.1 目的基因的克隆
重组木聚糖酶rXyn2441GH10的基因来自先前实验室保存的含有C.clariflavum的一个双催化域木聚糖酶基因Clocl-2441的重组质粒pET28a(+)-2441[18]。通过NCBI结构域序列分析将其中完整的GH10结构域进行引物设计,上游引物为:CCGGAATTCATTCCCTGCAGAAAATAATACAC,下游引物为:CCCAAGCTTCATTAATATTTCTTTCAATGCGTTG,扩增产物含有1 062 bp,表达蛋白含有353个氨基酸。以pET28a(+)-2441重组质粒为模板,利用PrimeSTAR MaxDNA Polymerase高保真DNA聚合酶扩增出完整目的基因片段。PCR反应条件为95 ℃预变性5 min,98 ℃变性10 s,55 ℃退火10 s,72 ℃延伸90 s,34个循环,最后72 ℃延伸10 min。
1.2.2 重组载体pET28a-Xyn2441GH10的构建
将保藏的含有pET28a(+)质粒的JM109菌株培养12~14 h后提取质粒。将扩增的目的产物片段和空载质粒用限制性内切酶EcoR I、Hind III在37 ℃下反应1 h,将双酶切产物胶回收后测定核酸含量和纯度。把载体和目的片段以1∶8的比例用T4DNA连接酶过夜连接,将连接产物用热激法转化入JM109感受态细胞。转化后的重组子均匀涂布到含有100 μg/mL的卡那霉素固体LB培养基上,置于37 ℃培养箱中培养。挑取单菌落进行双酶切验证,将阳性重组质粒转化入大肠杆菌BL21(DE3)后测序验证。
1.2.3 重组木聚糖酶rXyn2441GH10的诱导表达及分离纯化
将构建好的重组菌于37 ℃、200 r/min摇床上过夜培养,以4%的体积分数接种量转接入含有100 μg/mL卡那霉素的LB液体培养基中,同样条件下培养至OD600达到0.6~0.8,向培养基中加入终浓度为0.4 mmol/L的异丙基-β-D-硫代吡喃半乳糖苷(isopropyl β-D-thiogalactoside,IPTG),25 ℃、200 r/min诱导表达10 h,离心收集菌体。将收集的菌体用磷酸缓冲液重新悬浮进行超声破碎,离心收集上清,粗酶液用0.22 μm水系滤膜过滤后通过1 mL镍柱进行纯化(GE公司his-TrapTMHP)。把有酶活性的洗脱峰通过脱盐柱(Sephadex G25)进行脱盐,十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(sodium dodecyl-sulfate-polyacrylamide gel electrophoresis, SDS-PAGE)电泳鉴定纯度,测定蛋白量和酶活。
1.2.4 重组木聚糖酶rXyn2441GH10的活性测定
将10 mg/mL的木聚糖底物与梯度稀释后的酶液反应10 min后,用3,5-二硝基水杨酸(dinitrosalicylic acid,DNS)法测定还原糖(以木糖计)[19]。通过标准曲线计算还原糖量和相应的酶活性(3个平行取平均值)。酶活单位定义为:每分钟水解木聚糖产生1 μmol还原糖所需的酶量为1 U。
1.2.5 重组木聚糖酶rXyn2441GH10的酶学性质
1.2.5.1 最适温度
在0.1 mol的磷酸缓冲溶液(pH 6.5)条件下,45~85 ℃范围内(梯度为5 ℃)检测酶活性,以最高酶活为100%计算相对酶活。
1.2.5.2 最适pH
在pH 5.0~8.0(梯度为0.5)缓冲溶液和最适温度条件下检测不同pH下的酶活,以最高酶活为100%计算相对酶活。
1.2.5.3 温度稳定性
将蛋白质量浓度为0.08 mg/mL的酶液,在55、60、65、70 ℃下处理不同的时间,在最适温度和pH下检测剩余酶活,以处理0 h的酶活为100%计算处理不同时间的剩余酶活。
1.2.5.4 pH稳定性
将酶液在pH 3.0~9.0范围的缓冲溶液中于20 ℃下处理5 h,在最适条件下测定其剩余酶活,以未处理的酶活为100%计算不同pH处理后剩余酶活。
1.2.5.5 金属离子及EDTA对重组木聚糖酶活性的影响
以Tris-HCl缓冲液配置不同金属离子母液,按需添加至稀释后的酶液中,分别考察了1 mmol/L和 5 mmol/L的 KCl、LiCl、CoCl2、MgCl2、MnCl2、CaCl2、FeCl3、AlCl3和EDTA 对rXyn2441GH10酶活的影响。在最适条件下检测酶活,以未添加金属离子和EDTA的酶活为100%计算酶活的变化。
1.2.6 重组木聚糖酶rXyn2441GH10的动力学参数测定
在酶的最适条件下测定不同底物浓度下的初始酶活,根据Lineweaver-Burk 双倒数作图。以底物1/[S]为X轴,1/[V]为Y轴作图拟合出线性方程,计算不同底物的Km和Vmax以及其他参数。
1.2.7 重组木聚糖酶GH10结构域209位点饱和突变
通过全质粒PCR来引入19种氨基酸突变,在引物上设计突变位点以重组质粒为模板通过高保真酶扩增质粒全长,DpnI酶消化PCR产物去除模板后转化入大肠杆菌BL21(DE3)感受态。挑选重组子测序确定正确的突变菌株。突变体菌株和rXyn2441GH10菌株在相同的条件(同1.2.3)诱导表达,控制菌体收集量一致,在相同的条件下测定rXyn2441GH10和突变酶的酶活。
2 结果与分析
2.1 目的基因的克隆与重组质粒的构建
以含有rXyn2441GH10木聚糖酶基因的质粒pET28a(+)-2441为模板进行PCR扩增,琼脂糖凝胶电泳验证PCR产物。电泳结果显示,目的基因条带单一,与理论1 062 bp大小符合(图1)。
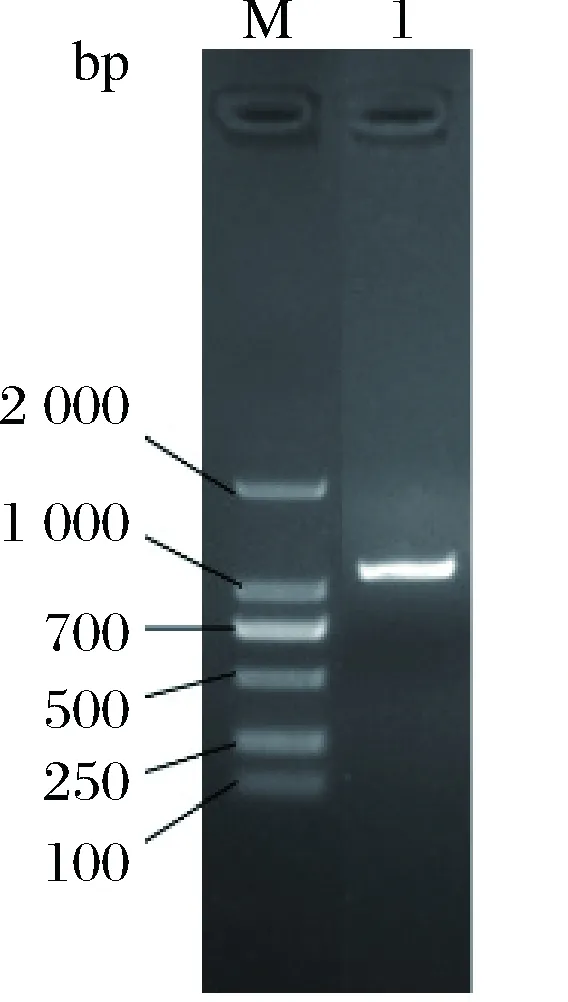
图1 目的基因Xyn2441GH10的PCR扩增Fig.1 PCR amplification of gene Xyn2441GH10
EcoR I和Hind III双酶切法将扩增的目的基因插入pET28a(+)载体上。将构建的重组质粒转化至E.coliBL21(DE3)中,双酶切重组质粒后通过电泳验证(图2)。
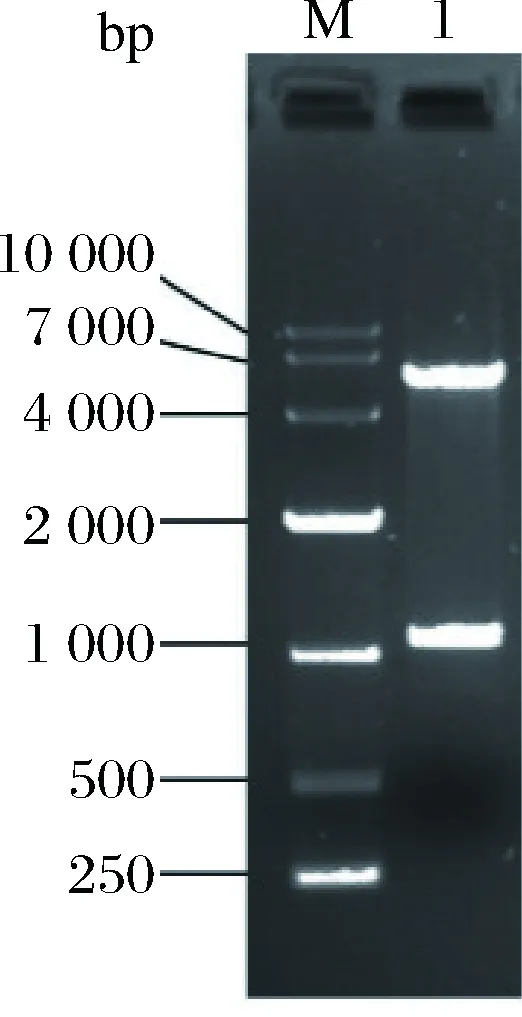
图2 重组质粒pET28a-Xyn2441GH10的双酶切验证Fig.2 Validation of recombinant plasmid by double enzyme digestion
1泳道有2条条带,第1条与载体大小符合;第2条与Xyn2441GH10基因理论大小1 062 bp符合,说明基因成功插入正确位点。将菌液样品送至华大基因测序。测序结果显示,插入的基因序列与美国国立生物技术信息中心(National Center for Biotechnology Information, NCBI)上公布的C.clariflavumDSM 19732基因Clocl-2441序列( ID:11562857)的GH10结构域基因序列完全一致,且读码框正确,表明重组质粒构建成功。将重组木聚糖酶的GH10结构域氨基酸序列与不同种属来源的GH10木聚糖酶氨基酸序列用DNAMAN比对分析,结果表明不同种属来源的GH10木聚糖酶氨基酸序列比较保守,相似度为64.01%。GH10木聚糖酶在N末端的氨基酸的保守性较低比较灵活,相比之下C末端和中间部分氨基端保守性较强(图3)。

1-Clostridium clariflavum; 2-Clostridium thermocellum; 3-Actinomadura meyerae; 4-Streptomyces olivaceoviridis; 5-Thermotoga maritime,黑色倒三角标示出推测的129位和240位催化氨基酸,黑色箭头标示出209位点的组氨酸图3 不同来源的GH10木聚糖酶氨基酸序列比对Fig.3 Amino acid sequence alignment of GH10 xylanase from different sources
2.2 重组木聚糖酶rXyn2441GH10的诱导表达及分离纯化
将重组E.coliBL21(pET28a-Xyn2441GH10)菌株进行诱导表达分离纯化(方法1.2.3)。把分别含有pET28a(+)的大肠杆菌、pET28a-Xyn2441GH10未诱导大肠杆菌、pET28a-Xyn2441GH10诱导后大肠杆菌上清酶液以及纯化后酶液,取相同蛋白量进行SDS-PAGE蛋白电泳。对比空质粒(1泳道)、重组质粒未诱导(2泳道)、重组质粒诱导(3泳道)发现:第3泳道在44.3 kDa处有明显的蛋白条带与重组木聚糖酶理论大小43.8 kDa符合,表明目的蛋白成功表达并且为可溶性蛋白(图4)。经过镍柱纯化和脱盐处理后rXyn2441GH10达到了电泳纯,可进行进一步的酶学性质分析。
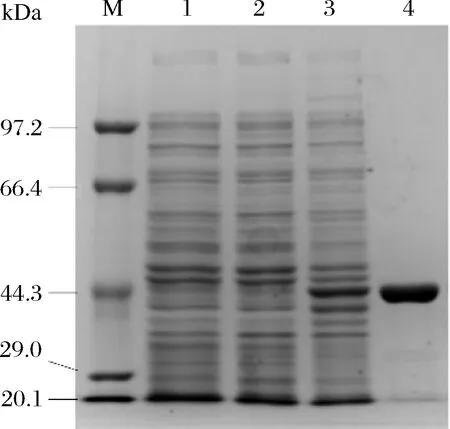
M-marker; 1-pET28a(+); 2-pET28a-Xyn2441GH10 (-IPTG); 3-pET28a-Xyn2441GH10 (+0.4 mmol/L IPTG); 4-pET28a-Xyn2441GH10 (纯化后)图4 重组木聚糖酶rXyn2441GH10的SDS-PAGE电泳Fig.4 SDS-PAGE of recombinant xylanase rXyn2441GH10
2.3 木聚糖酶rXyn2441GH10的酶学性质
2.3.1 最适温度
最适反应温度结果如图5所示。rXyn2441GH10的最适反应温度是70 ℃,与rXyn2441最适温度相同[18]。
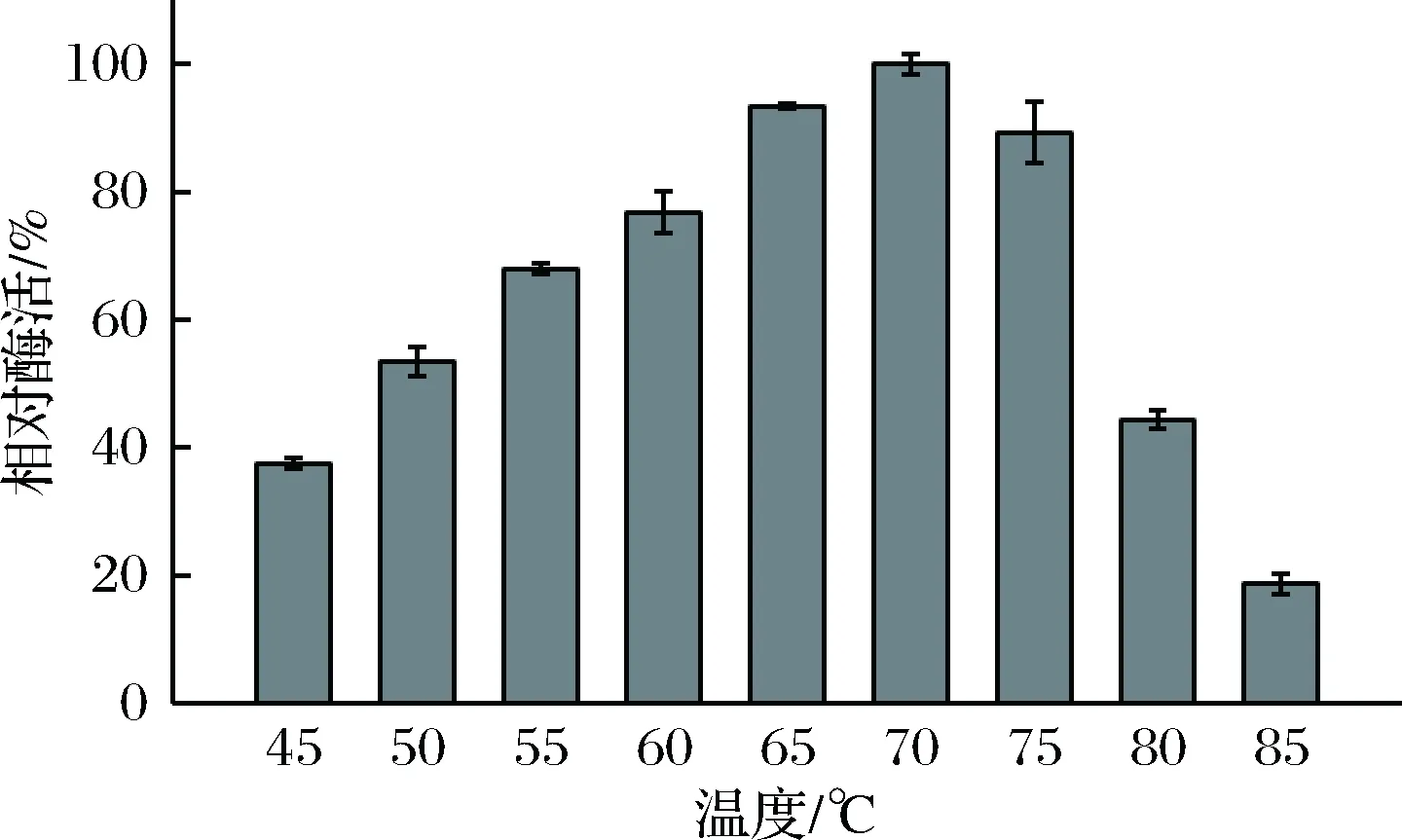
图5 重组木聚糖酶rXyn2441GH10的最适温度Fig.5 The optimal reaction temperature for recombinant xylanase rXyn2441GH10
rXyn2441GH10在60 ~75 ℃之间酶活比较高,75 ℃以上酶活急剧下降,85 ℃时酶活仅为70 ℃时的19%,rXyn2441GH10属于嗜热木聚糖。如表1所示,大多数GH10木聚糖酶最适温度在45 ~80 ℃之间,rXyn2441GH10木聚糖酶的最适温度较高,为在较高温度环境下的应用提供了新选择。
2.3.2 最适pH值
最适反应pH结果如图6所示。rXyn2441GH10的最适反应pH是7.0,在pH 6.0~7.5之间酶活为最适酶活的70%以上,在pH值小于5.5时其酶活下降较快,pH 5.0时的酶活仅为最适酶活的20%,属于中性木聚糖酶。与rXyn2441的最适pH相比提高了0.5,双结构域的最适pH不同可能是导致pH不同的原因[18]。不同来源的GH10木聚糖酶最适pH分布比较广泛,大多在pH 4.0~9.0之间,rXyn2441GH10适合在pH 6.0~7.5 的环境中应用(表1)。
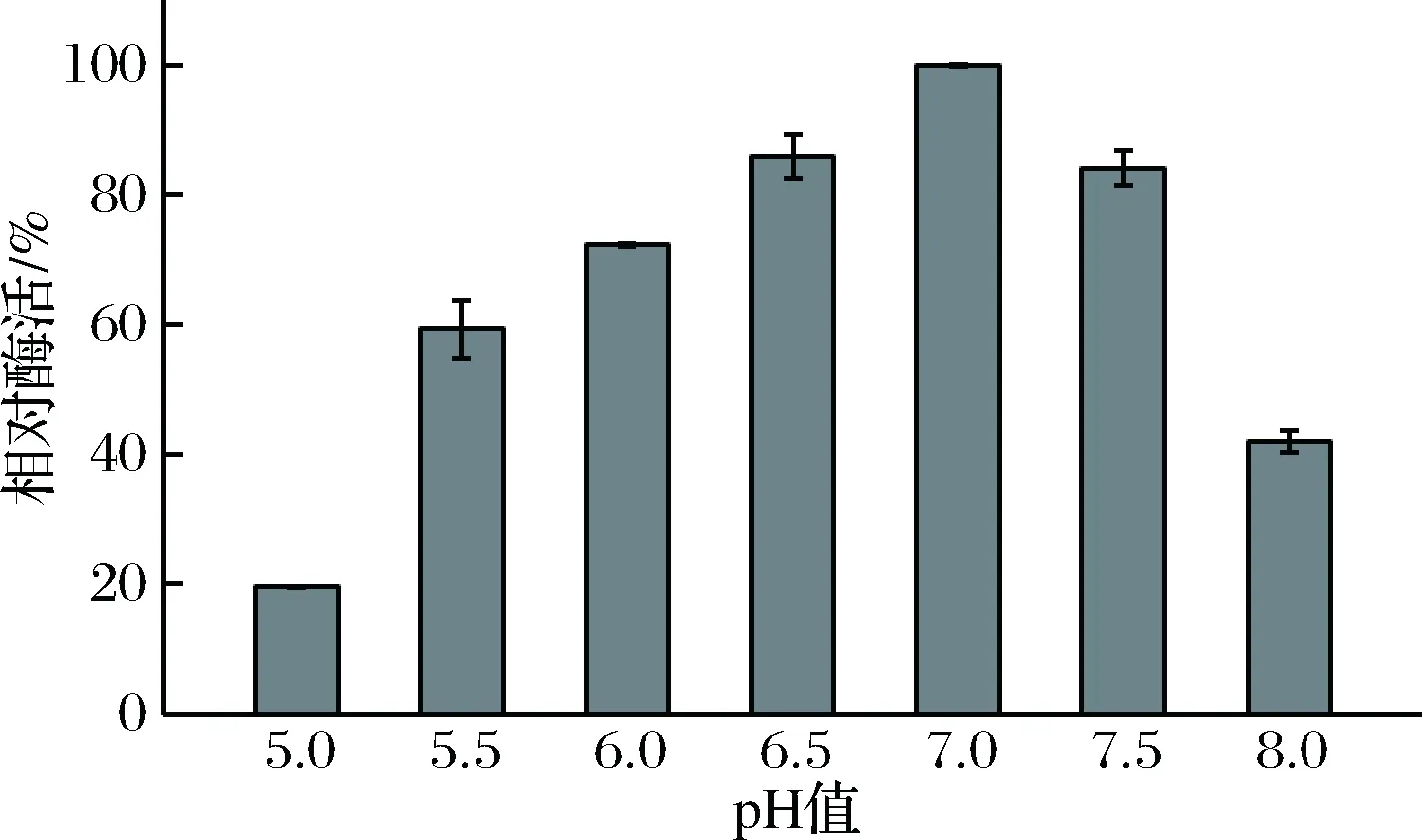
图6 重组木聚糖酶rXyn2441GH10的最适pHFig.6 The optimal reaction pH for recombinant xylanase rXyn2441GH10
2.3.3 温度稳定性
温度稳定性结果如图7所示。rXyn2441GH10在60 ℃及以下很稳定,在55、60 ℃处理6 h后剩余酶活为初始酶活的90%以上。rXyn2441GH10在65 ℃相比60 ℃稳定性有所下降,在处理6 h后酶活为初始酶活的38%。rXyn2441GH10在70 ℃下热稳定性明显降低,在处理1 h后酶活仅为初始酶活的17%。结果表明rXyn2441GH10在60 ℃及以下比较稳定,在70 ℃以及以上酶活稳定性降低。与多数不同来源的GH10木聚糖酶相比,rXyn2441GH10在65 ℃以下稳定性更强(表1)。
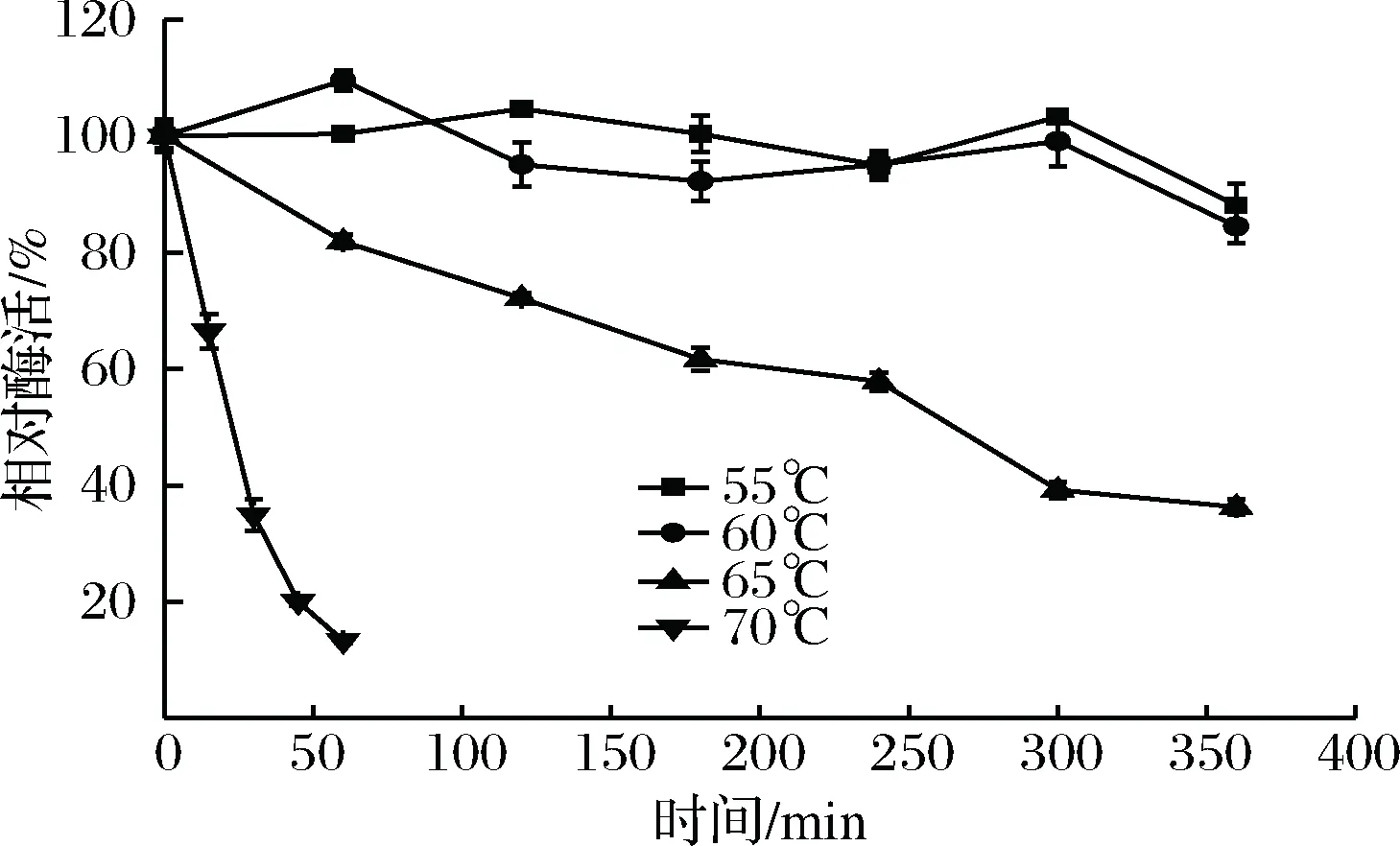
图7 重组木聚糖酶rXyn2441GH10的温度稳定性Fig.7 The thermostability for recombinant xylanase rXyn2441GH10
2.3.4 pH稳定性
pH稳定性结果如图8所示。rXyn2441GH10在pH 4.0~9.0之间都比较稳定,在该条件下处理5 h后其剩余酶活在84%以上。在pH 3.0环境下处理5 h后剩余酶活急剧下降仅为初始酶活的22%,有可能在该pH下对酶活性中心影响很大。rXyn2441GH10在中性偏酸性的环境下pH稳定性最好,与已知的GH10木聚糖酶相比pH稳定性范围更宽,适合的应用范围更广泛(表1)。
2.3.5 金属离子和EDTA对rXyn2441GH10的影响
结果如表2所示。K+、Ca2+、Mg2+、Al3+和Fe3+对rXyn2441GH10酶活有促进作用,其中5 mmol/L的Mg2+使的酶活提高了79.2%。Li+、Co2+和Mn2+对酶活有抑制作用,5 mmol的Co2+使酶活降低到58.3%。EDTA对酶活有微弱的促进作用。其中较高的金属离子浓度对rXyn2441GH10影响比较大,与NCBI数据表明蛋白存在较多金属离子结合位点一致[20],可能是金属离子与酶的结合进而影响了其的活性。以上结果对rXyn2441GH10的应用有重要的指导作用。
2.4 rXyn2441GH10的动力学参数
通过1.2.6方法测定数据用Origin 9.0作图拟合线性方程,榉木木聚糖底物拟合方程是Y=0.001 5X+0.000 591(R2=0.992),甘蔗渣木聚糖底物拟合方程是Y=0.001 6X+0.001 7(R2=0.992)。根据线性方程计算动力学参数(表3),相比之下重组酶水解榉木木聚糖效率更高,且与文献总结的不同来源的GH10木聚糖酶相比有更大的Vmax和kcat,但对羧甲基纤维素钠没有活性,与文献报道相一致[21-22]。
2.5 重组木聚糖酶209位点组氨酸的定点饱和突变
通过全质粒PCR引入突变的方法,将209位点的组氨酸突变成其他19种氨基酸。测序结果比对分析表明,209位点的组氨酸被成功突变成19种氨基酸。测定rXyn2441GH10和突变酶的酶活,以rXyn2441GH10酶活为100%作图,结果表明19个突变体对山毛榉木聚糖降解活性丧失或者几近丧失,活性最高的突变体H209S仅为rXyn2441GH10的0.17%(图9)。说明209位点的组氨酸对酶活性有很大的影响。

表3 重组木聚糖酶rXyn2441GH10动力学参数Table 3 The kinetic parameters of the recombinant xylanase rXyn2441GH10

图9 rXyn2441GH10的209位点饱和突变对酶活性的影响Fig.9 Effect of 209 site-saturation mutagenesis on activity of the recombinant xylanase rXyn2441GH10
为了研究突变体酶活变化是不是二级结构变化引起的,通过圆二色性光谱分析了H209N突变体和rXyn2441GH10的二级结构(图10)。
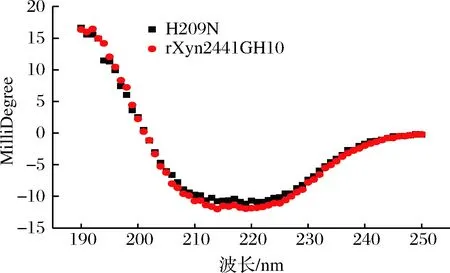
图10 重组木聚糖酶rXyn2441GH10和H209N突变酶的圆二色性光谱Fig.10 The circular dichromatic spectrum of recombinant xylanase rXyn2441GH10 and its H209N mutant
结果表明,H209N突变体和rXyn2441GH10在二级结构上没有发生变化,突变后酶活的变化不是二级结构变化引起的。将不同来源的GH10木聚糖酶的氨基酸序列比对分析(图3),箭头标注的组氨酸和倒三角标注的2个谷氨酸在所有GH10结构域中非常保守,且这3个氨基酸周围的氨基酸同样很保守。rXyn2441GH10与同属菌Clostridiumthermocellum来源的GH10木聚糖酶XynZ氨基酸序列相似度为63%,且XynZ晶体结构已经解析。在XynZ中2个保守的谷氨酸为催化氨基酸,209位点的组氨酸在结构中位于酶的催化裂口处,与2个催化氨基酸Glu相邻可与催化残基产生氢键作用,使得2个谷氨酸在空间结构保持异头构象完成催化[23]。在rXyn2441GH10中209位点的His有可能起到类似的作用,209位点突变成其他的氨基酸后导致异头构象不稳定使得催化效率大大降低甚至丧失。
3 结论
Clostridiumclariflavum菌株可产生多种木质纤维素降解酶类,在木质纤维素降解方面有广泛的应用潜力。目前国外对C.clariflavum菌株来源的木质纤维素降解酶类主要关注在以纤维小体为中心的复合酶的作用机制以及应用,但对单个酶或单个结构域性质研究很少,对工业应用指导作用有限[18,24]。本实验成功克隆了Clocl-2441上GH10结构域木聚糖酶基因并在大肠杆菌中表达为可溶性蛋白。酶学性质研究表明该酶的最适反应温度为70 ℃,最适pH值为7.0,属于中性嗜热木聚糖酶。rXyn2441GH10适合在55~65 ℃、pH 6.0~7.5条件下应用,与已报道的部分GH10木聚糖酶相比具有更广泛的应用适应性。209位点的组氨酸是rXyn2441GH10活性的关键氨基酸,并且在GH10结构域中极其保守,对GH10木聚糖酶的位点分析和改造有指导意义。本研究结果为重组GH10木聚糖酶的研究及工业应用提供了一定的理论依据。
