绵羊弓形虫TgSheepHn1对小鼠的病理损伤及其抗原分布特点
2018-06-21,,,,,
,,, ,,
刚地弓形虫(Toxoplasmagondii)是一种细胞内寄生原虫,广泛分布于全球陆地、水域范围内,能够侵染所有恒温动物的有核细胞,从而诱发弓形虫病[1]。猫科动物是弓形虫的终末宿主,绵羊可经过食入含卵囊的饲草或饮水、孕期垂直传播胎儿而感染弓形虫[2],弓形虫病是诱发绵羊流产的病原之一(www.gov.uk/government/statistics)。研究发现,人类和绵羊妊娠期首次感染弓形虫,易引发胎儿发育异常或流产[3],因此弓形虫病对绵羊产业以及人类健康造成危害。分离弓形虫虫株并研究其致病特征是防控弓形虫病的基础工作,目前我国从绵羊体内共分离到3株弓形虫虫株[4-5],本文研究了绵羊分离株TgSheepHn1(ToxoDB#9)[5]对Swiss小鼠的病理损伤及抗原分布特点,补充了我国绵羊分离株的致病特点,为进一步了解弓形虫病的基本特征提供相关依据。
1 材料与方法
1.1弓形虫虫株速殖子与抗体 TgSheepHn1(ToxoDB#9)弓形虫虫株分离于河南省焦作市绵羊心肌[5],虫株经Vero细胞培养获得大量速殖子,保存于本研究室。兔抗弓形虫多克隆抗体、弓形虫抗原均由美国农业部Dubey实验室馈赠,小鼠Specific HRP/DAB(ABC)Detection IHC kit 购自abcam公司(货号:ab64264)。
1.2实验动物及处理 Swiss小鼠15只,≥45 d龄,体重≥25 g,弓形虫检测阴性,购于河南省实验动物中心。小鼠分为3组,2个感染组,1个对照组,每组5只小鼠,雌雄分开,自由采食清洁级全营养颗粒饲料,饮用高压灭菌水。感染组小鼠分别腹腔注射104浓度的弓形虫速殖子,对照组小鼠注射无菌生理盐水,接种量均为1 mL/只。每日观察小鼠临床表现并记录死亡情况。对感染后死亡小鼠的肺脏及肠系膜淋巴结涂片镜检弓形虫速殖子,并用10%的中性福尔马林溶液固定小鼠大脑、舌头、眼球、心脏、肝脏、脾脏、肺脏、肾脏、肾上腺、小肠、肠系膜淋巴结、生殖器官、膀胱和骨骼肌。
1.3实验方法 肺脏及肠系膜淋巴结中弓形虫速殖子检测采用组织涂片方法,光学显微镜镜检观察;运用常规病理学技术制作石蜡切片,采用苏木素与伊红对比染色法(Hematoxylin-Eosin staining,H&E)鉴定病理损伤,免疫组织化学(Immunohistochemistry,IHC)染色,定位弓形虫抗原分布。
2 结 果
2.1Swiss小鼠接种TgSheepHn1弓形虫速殖子后的发病情况 腹腔接种104浓度的TgSheepHn1弓形虫速殖子后,感染小鼠在72 h开始出现被毛逆立、弓背蜷缩、呼吸窘迫和食欲降低等临床表现,感染组小鼠在接种后10 d(10 days post inoculation,10 DPI)内100%死亡,最早急性死亡时间为8 DPI,对照组小鼠未见异常(表1)。对急性期死亡小鼠肺脏和肠系膜淋巴结涂片镜检,可见大量弓形虫速殖子,见图1(A、B),确定小鼠因弓形虫感染而死亡。
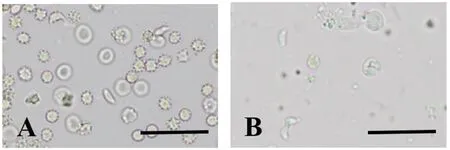
A: Tachyzoites of T. gondii, lung smear, unstained, 8 DPI; B: Tachyzoites of T. gondii, mesenteric lymph nodes smear, unstained, 8 DPI. Bar = 50 μm图1 小鼠组织涂片的弓形虫速殖子Fig.1 Tachyzoites of T. gondii in tissue smear of mice
表1 Swiss小鼠腹腔注射104浓度的TgSheepHn1弓形虫速殖子的发病情况
Tab.1 Pathogenicity of the Swiss mice post inoculation with 104of TgSheepHn1 T. gondii tachyzoites by intraperitoneal injection
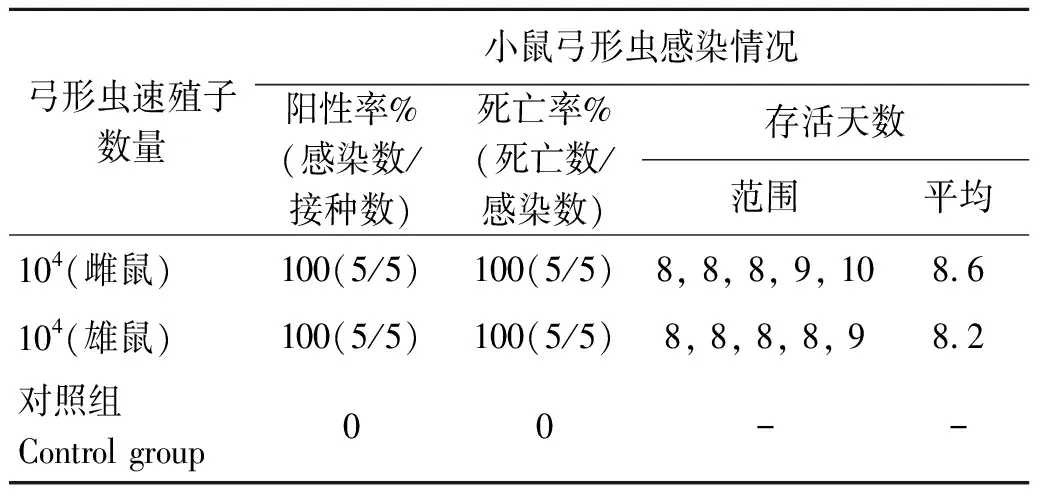
弓形虫速殖子数量小鼠弓形虫感染情况阳性率%(感染数/接种数)死亡率%(死亡数/感染数)存活天数范围平均104 (雌鼠)100(5/5)100(5/5)8, 8, 8, 9, 108.6104 (雄鼠)100(5/5)100(5/5)8, 8, 8, 8, 98.2对照组Control group00--
注:“-”表示小鼠未感染
2.2感染TgSheepHn1弓形虫小鼠的病理组织损伤及弓形虫抗原分布 死亡小鼠剖检主要眼观病变为:肺脏暗红色,淤血,脾脏和肠系膜淋巴结肿大,肝脏肿大淤血,肾上腺肿大,小肠肠壁菲薄充血。对各组织器官做常规石蜡切片,H&E染色镜检发现,多器官表现为坏死、水肿,血管周围炎,但未见出血。急性死亡小鼠组织器官内弓形虫及其分泌蛋白等抗原分布于多处组织,其中肝脏胆囊管肌层、脾脏、肠系膜淋巴结、肾上腺、生殖系统有大量分布。肝脏肝细胞呈局灶性坏死,血管周围炎性细胞浸润,肝脏胆囊管肌层大量单核细胞浸润,可见大量弓形虫虫体的断面分布(图2A,图3A);肠系膜淋巴结大量淋巴细胞坏死、水肿,分布大量弓形虫虫体(图2B,图3B);脾脏白髓坏死(图2C,图3C);回肠肠黏膜上皮细胞坏死脱落崩解,肠腺萎缩(图2D);肺脏肺泡间隔增宽,单核细胞浸润(图2E);肾小管上皮脱落坏死,单核细胞浸润(图2F);肾上腺固有结构不清晰,可见局灶性细胞坏死(图2G)及大量弓形虫虫体(图3D);大脑和心肌未见明显病理损伤,但可见弓形虫抗原分布(图3 E、F);睾丸、附睾和前列腺可见局灶性坏死,分布大量弓形虫虫体(图2H,图3G、H);子宫固有结构破坏,弓形虫分布于黏膜层、黏膜下层、肌层及浆膜层(图2I,图3I)。此外,肺脏、肾脏、小肠、膀胱及尿液中均可见弓形虫抗原分布,有的可以看到假包囊。从弓形虫抗原分布规律,表现为各脏器均是由外膜到中心逐渐减少。
3 讨 论
弓形虫虫株毒力分析研究表明:Ⅰ型为强毒株,感染性强,急性死亡率高;Ⅱ、Ⅲ型和非典型虫株均属于弱毒株,致死率低,多为高成囊率,常常诱发慢性弓形虫病,但在试验条件下,高浓度的弱毒株对小鼠也具有较高的致死率,如对远交小鼠、近交小鼠和免疫缺陷小鼠[6-7]。对比原型克隆谱系典型参考虫株和我国已分离的其他动物源的ToxoDB#9型虫株,本试验所研究的绵羊分离株TgSheepHn1(ToxoDB#9)弓形虫对Swiss小鼠的感染性和致死性均较强,与已报道的我国优势基因型虫株ToxoDB#9型(TgCtgy1、TgCtwh3和TgCtwh6等虫株)虫株的侵染毒性特征相似[5,8]。
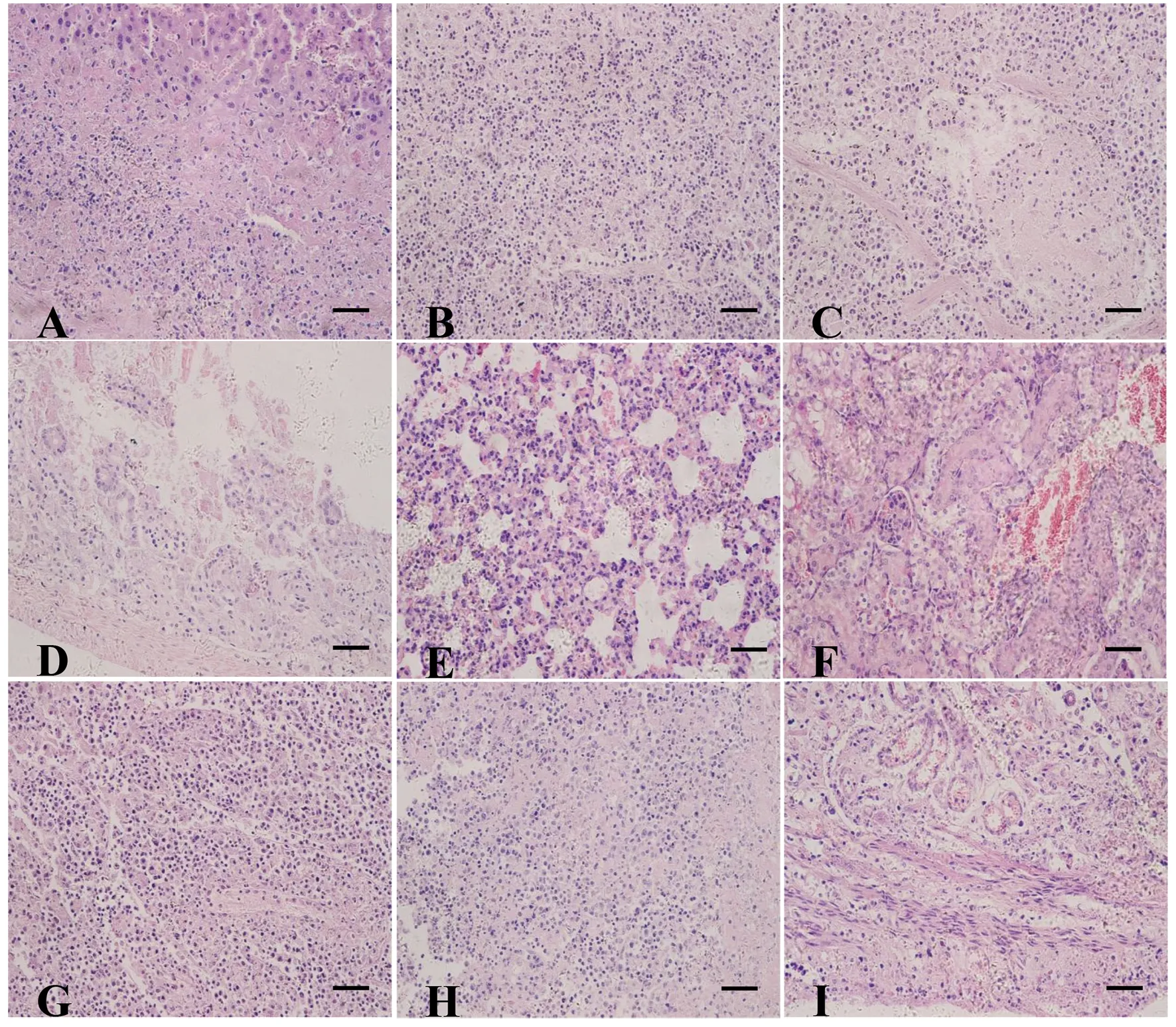
A-I are liver, mesenteric lymph nodes, spleen, ileum, lung, kidney, adrenal, epididymis, uterus, respectively. HE staining, bar=50 μm.图2 Swiss小鼠感染TgSheepHn1弓形虫后,各器官的病理组织损伤, 8 DPI.Fig.2 Pathology histological lesions of Swiss mice after infection with TgSheepHn1 tachyzoites, 8 DPI

A-I are liver, mesenteric lymph nodes, spleen, adrenal, brain, heart, testis, prostate, uterus, respectively. IHC staining, bar=50 μm.图3 Swiss小鼠感染TgSheepHn1弓形虫后,各器官弓形虫抗原的分布, 8 DPI.Fig.3 Antigens distribution of T. gondii in tissue of Swiss mice after infection TgSheepHn1 tachyzoites, 8 DPI
腹腔接种104浓度的TgSheepHn1弓形虫速殖子后,小鼠在感染后约8 d死亡,对其病理损伤及虫体抗原分布分析发现,大部分组织器官均表现不同程度的损伤,其中肺脏、脾脏、肠系膜淋巴结、回肠、生殖器官,肾上腺损伤较重,以组织细胞坏死,水肿为特征,且这些脏器弓形虫抗原分布量较多。这与已报道的Ⅰ型强毒株CT1型弓形虫不同浓度卵囊灌胃远交系昆明小鼠后,104个卵囊即可导致小鼠在7 DPI时全部死亡,且急性死亡小鼠的各组织脏器损伤程度及抗原分布趋势基本相同[9]。研究发现,108个VEG株弓形虫的卵囊可导致昆明小鼠100%死亡,而从猫组织分离到的ToxoDB#17(TgCatCHn2)型和ToxoDB#9(TgCatCHn4)型弓形虫,1个卵囊可引起小鼠感染,但随着卵囊浓度的增加并不会引起小鼠死亡[10-13]。对比已研究的报道可推断,TgSheepHn1弓形虫对Swiss小鼠的毒力表现较强,介于强毒和弱毒之间。
对BALB/c小鼠的急性弓形虫病理组织损伤研究发现,≥105浓度的VEG、ME49和GT1株弓形虫卵囊对小鼠在4~10 d内急性感染期的组织损伤程度依次为肠系膜淋巴结、小肠、脾脏、肺脏和肝脏,大部分组织均出现损伤,其中回肠段损伤最重,回肠为卵囊灌胃弓形虫的主要入侵门户[7]。此绵羊分离株TgSheepHn1弓形虫速殖子在急性感染期对昆明小鼠的各脏器侵染性较强,损伤与研究报道的也基本一致,但十二指肠、空肠和回肠的病理损伤程度无差异,推测腹腔注射弓形虫速殖子对整个小肠肠道几乎同时入侵,这种差异可能与接种方式和接种虫体的状态有关,也可能是虫体固有毒力差异性的表现,具体原因还需验证。另外,急性期感染弓形虫对小鼠小肠潘氏细胞的破坏会引起溶菌酶表达缺失,强毒株ToxoDB#216型表现为肠隐窝损伤严重,弱毒株ToxoDB#17型、ME49株弓形虫表现为肠隐窝轻度损伤,溶菌酶表达缺失[14-16]。TgSheepHn1弓形虫可引起回肠肠腺萎缩,是否会引起溶菌酶缺失现象尚待进一步的研究。
本研究发现,急性期死亡小鼠多器官分布弓形虫速殖子及其分泌蛋白抗原,其中脾脏、肾上腺、肠系膜淋巴结、整个小肠和生殖系统分布密度最高,其余组织如肺脏、肝脏、肾脏、心肌和大脑也可观察到弓形虫抗原分布,各脏器外膜抗原分布量高推测与腹腔接种弓形虫速殖子有关,通过各脏器外膜的疏松结缔组织侵入各器官。急性死亡期小鼠生殖系统和肾上腺的弓形虫抗原分布较多,提示该虫株具有潜在的嗜生殖系统能力(包括雌性和雄性),且已有报道绵羊弓形虫可以通过精液传播[17-18],但关于弓形虫嗜肾上腺组织细胞和生殖系统间质细胞的能力未见相关研究报道。在研究绵羊分离株TgSheepHn1(ToxoDB#9)型弓形虫对昆明小鼠的病理损伤以及抗原分布过程中,发现的这些现象还需要我们进一步深入探讨,本报道为中国虫株对动物机体的病理损伤及抗原分布特点做出补充。
[1] Dubey JP. Toxoplasmosis of animals and humans[M]. Boca Raton: CRC Press, Taylor & Francis Group, 2010: 1-313.
[2] Innes EA. A brief history and overview ofToxoplasmagondii[J]. Zoonoses Public Health, 2010, 57 (1): 1-7. DOI: 10.1111/j.1863-2378.2009.01276.x
[3] Innes EA, Bartley PM, Buxton D, et al. Ovine toxoplasmosis[J]. Parasitology, 2009, 136 (14): 1887-1894. DOI: 10.1017/S0031182009991636
[4] Zhou P, Zhang H, Lin RQ, et al. Genetic characterization ofToxoplasmagondiiisolates from China[J]. Parasitol Int, 2009, 58 (2): 193-195. DOI: 10.1016/j.parint.2009.01.006
[5] Yang YR, Feng YJ, Yao QX, et al. Seroprevalence, isolation, genotyping, and pathogenicity ofToxoplasmagondiistrains from sheep in China[J]. Front Microbiol, 2017, 8: 136. DOI: 10.3389/fmicb.2017.00136
[6] Wang L, Chen H, Liu D, et al. Genotypes and mouse virulence ofToxoplasmagondiiisolates from animals and humans in China[J]. PLoS One, 2013, 8 (1): e53483. DOI: 10.1371/journal.pone.0053483
[7] Dubey JP, Ferreira LR, Martins J, et al. Oral oocyst-induced mouse model of toxoplasmosis: effect of infection withToxoplasmagondiistrains of different genotypes, dose, and mouse strains (transgenic, out-bred, in-bred) on pathogenesis and mortality[J]. Parasitology, 2012, 139 (1): 1-13. DOI: 10.1017/S0031182011001673
[8] Li M, Mo XW, Wang L, et al. Phylogeny and virulence divergency analyses ofToxoplasmagondiiisolates from China[J]. Parasit Vectors, 2014, 7 (1): 133-144. DOI: 10.1186/1756-3305-7-133
[9] 付晓莹, 孔阳光, 梁宏德, 等. 昆明小鼠感染CT1型弓形虫卵囊的虫体分布及病理组织学研究 [J]. 中国细胞生物学学报,2015, 37(10): 1377-1380.
[10] 陆瑶瑶, 冯永杰, 董辉, 等. VEG株弓形虫卵囊对昆明小鼠的毒力研究 [J]. 中国人兽共患病学报,2017, 33(7): 624-627. DOI:10.3969/j.issn.1002-2694.2017.07.010
[11] Yang YR, Ying YQ, Verma SK, et al. Isolation and genetic characterization of viableToxoplasmagondiifrom tissues and feces of cats from the central region of China[J]. Vet Parasitol, 2015, 211 (3-4): 283-288. DOI: 10.1016/j.vetpar.2015.05.006
[12] 王凯, 冯永杰, 付晓莹, 等. ToxoDB#17型弓形虫卵囊对昆明小鼠致病性的研究 [J]. 中国人兽共患病学报,2017, 33(1): 49-52. DOI:10.3969/j.issn.1002-2694.2017.01.009
[13] Yang YR, Feng YJ, Lu YY, et al. Antibody detection, isolation, genotyping, and virulence ofToxoplasmagondiiin captive felids from China[J]. Front Microbiol, 2017, 8: 1414. DOI: 10.3389/fmicb.2017.01414
[14] 王凯, 陆瑶瑶, 刘振阳, 等. ToxoDB#17型弓形虫感染小鼠引起小肠潘氏细胞溶菌酶缺失[J]. 中国细胞生物学学报,2017, 39 (2): 1-6.
[15] 张琼方, 付晓莹, 王凯, 等. 潘氏细胞在昆明小鼠弓形虫病发病过程中的变化[J]. 中国细胞生物学学报,2015, 37 (10): 1381-1385.
[16] Raetz M, Hwang SH, Wilhelm CL, et al. Parasite-induced TH1 cells and intestinal dysbiosis cooperate in IFN-gamma-dependent elimination of Paneth cells[J]. Nat Immunol, 2013, 14 (2):136-142. DOI: 10.1038/ni.2508
[17] Consalter A, Silva AF, Frazao-Teixeira E, et al.Toxoplasmagondiitransmission by artificial insemination in sheep with experimentally contaminated frozen semen[J]. Theriogenology, 2017, 90: 169-174. DOI: 10.1016/j.theriogenology.2016.12.004
[18] Bezerra MJ, Cruz JA, Kung ES, et al. Detection ofToxoplasmagondiiDNA in fresh and frozen semen from rams in Brazil[J]. Reprod Dom Anim, 2014, 49 (5): 753-755. DOI: 10.1111/rda.12361
