有氧运动对雄性肥胖大鼠生殖功能及睾丸组织TNF-α、Fas和Caspase-3 mRNA表达的影响
2018-06-20郑锡明孙玉霞姚婷婷衣雪洁
常 波,郑锡明,李 晶,高 畅,孙玉霞,姚婷婷,李 歌,衣雪洁
(1.沈阳体育学院国家体育总局重点实验室,辽宁 沈阳 110102;2.沈阳体育学院运动人体科学学院,辽宁 沈阳 110102;3.沈阳体育学院研究生部,辽宁 沈阳 110102)
近年来,国内外患有生育障碍的患者数目急剧升高,其中肥胖导致的生殖功能低下的现象已引起大家的广泛关注。最近对人类和其他物种的研究表明,长期肥胖会使雄性性腺机能减退(secondary hypogonadotropic),表现为雄性激素分泌不足、雌激素水平升高、精子质量参数下降和生殖功能出现障碍[1-2],但其作用的机制尚不清楚。有研究显示肥胖引起的雄性生殖功能障碍与生精细胞凋亡关系密切,雄性动物睾丸内生精细胞凋亡是正常的生理状态[2],但由于一些特殊的理化因素导致其大量凋亡,直接影响精子的生成数量和质量。调控细胞凋亡的主要途径有3种:死亡受体途径、线粒体途径和内质网途径。半胱氨酸蛋白酶(Caspase-3)是介导细胞凋亡的一类蛋白水解酶,是细胞凋亡过程中的一个关键酶,在细胞凋亡中起着不可替代的作用,参与了细胞凋亡的诱导,是CTL细胞杀伤机制的重要组成部分。作为死亡受体途径中最后的公共凋亡效应因子,可引发生殖细胞程序性死亡,一旦被大量激活,凋亡就不可避免。研究发现肥胖可影响睾丸组织中Caspase-3的水平,进而导致生殖细胞大量凋亡。
运动作为目前公认的治疗肥胖最为有效的手段之一,对雄性激素的影响还有很大的争议,运动形式、运动量和运动强度的差异可能会带来不同的实验结果。有关有氧运动对缓解肥胖个体的生殖细胞凋亡的影响报道还较少,进一步研究引发的凋亡机制则更少。为此,通过施加有氧运动干预改善大鼠的肥胖状态,并通过中枢和外周两条途径探讨肥胖状态下有氧运动对雄性生殖机能作用机制,力图为揭示有氧运动影响精子质量的机制提供实验依据,也为探究防治肥胖引起的生殖机能低下的有效途径提供理论参考。
1 研究对象与方法
1.1 研究对象
24只5周龄雄性SD大鼠,购于北京大学医学部。依据北京大学动物实验室动物伦理标准监督执行。饲养温度为(22±2)℃,分笼饲养,自由饮水。
1.2 实验方法
1.2.1 肥胖模型的制备
24只SD雄性大鼠随机分成2组,各12只,即正常饮食组(普通饲料)和高脂饮食组(高脂饲料)。分别为大鼠提供普通饲料和高脂饲料,饲养12周。高脂饮食组肥胖鼠建模成功后,将正常饮食组和肥胖(高脂饮食)组大鼠分别按照体重分层抽样再各自分为两组,即正常对照组(NC,6只)、正常饲料运动组(NE,6只)和肥胖组(OC,6只)、肥胖运动组(OE,6只)。高脂组大鼠的体重均在正常组大鼠平均体重的120%以上,肥胖建模成功[3]。
1.2.2 分组运动干预
两组运动组(正常运动组和肥胖运动组)共进行为期8周的跑台运动。正式运动开始前进行为期一周的适应性训练,以减少正式实验时的应激反应。起始时间为20 min,跑台速度设定为26 m/min,以后每天增加10 min,逐渐增加到60 min/d为止。保持这一负荷进行8周跑台运动,每周休息1 d。
1.2.3 样品的取材及处理
8周运动干预结束后,在末次运动后36~40 h取材,取材前禁食12 h。麻醉处死,迅速取两侧睾丸,用生理盐水冲洗后用滤纸吸干,转入液氮冷冻,以备实验使用。
1.2.4 测试指标及方法
1.2.4.1 睾丸组织重量、睾丸系数与精子活率测定
1)将大鼠的双侧睾丸从阴囊中分离提取出来,电子天平上称量:睾丸系数(%)=(睾丸重量/体重)×100%。2)动物处死后,迅速取一侧附睾置于37℃生理盐水2 mL小烧杯中,剪碎,再加入同等浓度的盐水,并与生理盐水以1比1的比例混合,经滤纸过滤后,提取滤液1 mL,加入生理常温下的生理盐水9 mL,混匀备用。取精子悬液中一滴放置于载玻片上,加盖玻片后,光镜观察精子的活动能力。依照世界卫生组织,观察精子的流动状态与形式,可把精子的活动度大致分为4个级别:1级:精子可以保持直线运动水平并且活动踊跃,观察到活动性优良。2级:基本可以保持直线运动,但与1级比较活动性略差。3级:精子保持直接向前运动的能力弱,只能在原处活动或旋转运动。4级:精子无任何运动能力。实验的每只大鼠都需要检查大概200个精子,然后计算精子的活率:精子活率=(1+2+3级精子数/1+2+3+4级精子数)×100%。
1.2.4.2 组织提取和RT-PCR检测 1)样品RNA的提取。①将睾丸样品从-80℃冰箱中取出,马上在天平上进行称重。然后在正常室温条件下放置5 min,待其充分溶解(将实验中所用的镊子、剪刀、匀浆器均进行无菌消毒后用DEPC水处理)。②分离RNA。③低温离心。④RNA沉淀。⑤清洗RNA。⑥干燥RNA。⑦溶解RNA沉淀。2)检测样品浓度,使用微量分光光度计。3)逆转录反应。依据Hiscript®IIOne Step qRT-PCR SYBR®Green试剂盒说明书,严格按照说明书完成整个设定步骤的实验工作。4)引物序列:实验中扩增引物序列,从PubMed数据库查询得到。引物具体序列见表1。

表1 RT-PCR反应相关引物的序列
1.2.5 统计学处理
使用SPSS 17.0统计软件进行数据分析,均数±标准差±S)表示实验结果。组间均数比较采用单因素方差分析的Scheffe’s Post hoc comparisons test。OC、OE组采用双因素方差分析来阐明运动和饮食干预对雄性肥胖大鼠生殖功能及睾丸组织基因表达的交互作用,检验标准:P<0.05为组间差异显著,P<0.01为组间差异极显著。
2 研究结果
2.1 各组大鼠体重、脂体比、睾丸重量、睾丸系数
由表2可知,通过8周的有氧运动,大鼠的肥胖情况有所减轻,睾丸系数上升,说明有氧运动可以减重,缓解了肥胖对睾丸形态结构的影响,同时有氧运动对于正常状态下机体的睾丸形态结构也有明显的改善作用。
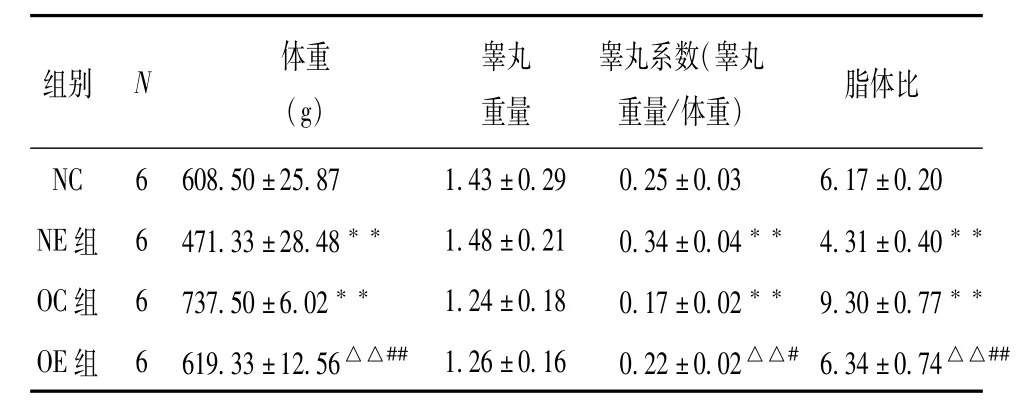
表2 各组大鼠体重、睾丸重量、睾丸系数、脂体比情况(¯X±S)
2.2 各组大鼠精子质量参数
如表3所示,经过20周高脂膳食喂养后的肥胖大鼠精子活性与精子数量明显下降,说明肥胖会对生殖功能造成影响。8周有氧运动能够较好地增强与改善正常和肥胖大鼠的精子活性,提高生殖能力。

表3 各组大鼠精子活动率的变化情况(¯X±S)
2.3 各组大鼠睾丸组织中的TNF-α、Fas、Caspase-3 mRNA的表达
如表4和图1所示,与正常组的大鼠相比,正常运动组的大鼠睾丸中TNF-α、Fas、Caspase-3的基因转录水平均显著性降低(P<0.05);肥胖组大鼠睾丸组织中的TNF-α、Fas、Caspase-3的基因转录水平均显著性升高(P<0.05、P<0.01、P<0.01)。同肥胖组的大鼠相比较,肥胖运动组的大鼠睾丸组织中的TNF-α、Fas、Caspase-3基因转录水平均明显降低(P<0.01、P<0.05、P<0.01)。同正常运动组相比,肥胖运动组的大鼠睾丸组织中的TNF-α、Fas、Caspase-3基因转录水平显著性提升(P<0.01、P<0.01、P<0.05)。
表4 各组大鼠睾丸组织TNF-α、Fas、Caspase-3 mRNA表达水平(±S)
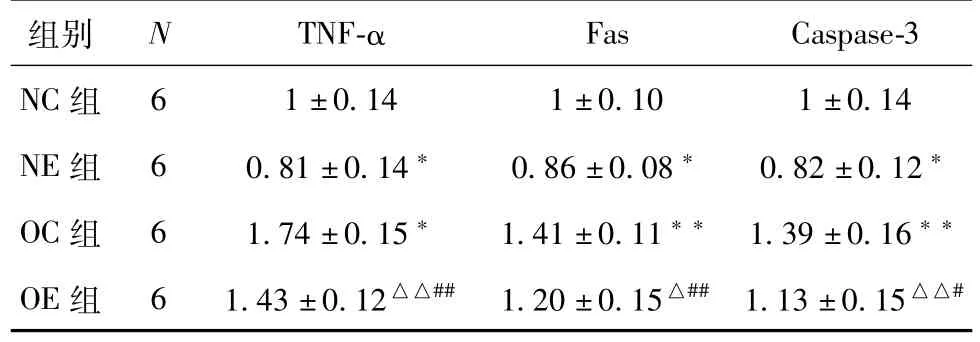
表4 各组大鼠睾丸组织TNF-α、Fas、Caspase-3 mRNA表达水平(±S)
Fas Caspase-3 NC组组别N TNF-α 6 1±0.14 1±0.10 1±0.14 NE组6 0.81±0.14*0.86±0.08*0.82±0.12*OC组6 1.74±0.15*1.41±0.11**1.39±0.16**OE组6 1.43±0.12△△##1.20±0.15△##1.13±0.15△△#
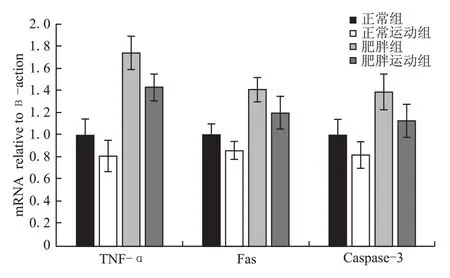
图1 TNF-α、Fas、Caspase-3 mRNA表达水平
3 分析与讨论
3.1 有氧运动对肥胖大鼠生殖功能的影响
睾丸作为雄性动物最重要的生殖器官,其发育与成熟状况将对雄性生殖能力产生很大的影响[4-9]。本实验发现肥胖组大鼠体重、脂体比明显提高,前期研究发现肥胖组体内血清睾酮、黄体生成素与卵泡刺激素水平明显降低,雌二醇明显升高[3];睾丸系数、精子数目和精子活动率显著降低;而肥胖运动组大鼠的体重、脂体比明显下降。前期研究还发现血清睾酮、黄体生成素与卵泡刺激素的含量有显著性升高,雌二醇明显下降[3];睾丸系数、精子数目和精子活动率均显著上升,提示运动可改善睾丸的形态结构。运动作为一种减脂的手段,不仅能通过增加能量消耗和基础代谢来降低体重,可能还会因此而改善睾丸功能。
李玲[10]在研究中发现肥胖使大鼠生精细胞大量下降且排列紊乱、结构异常,同时还会影响生精功能,精子异常比率显著高于普通组。徐庆阳[9]研究发现肥胖使精子数目与活动力均有下降的现象。临床研究也发现肥胖人群存在精子数目和质量下降、生精功能受损等现象[12]。肥胖造成精子质量下降的主要原因,在于肥胖会影响下丘脑—睾丸—性腺轴(HPT)的功能,长期的高热量摄入使得雄性机体的脂肪组织大量聚集,过量堆积的脂肪能够通过芳香化作用使雄性激素雌性化,导致雌激素含量上升,反馈性抑制HPT轴的功能,造成生精障碍。同时体内精子的发生与睾酮有着重要的关系。在生精小管内维持适量的睾酮水平能够有助于精子的产生与成熟。有研究表明[6],随着BMI的上升,男性血清睾酮的水平有下降的趋势,同时精子密度也相应减少。
运动对机体是一种应激刺激,会引起一系列神经-内分泌的改变。有研究发现[9],运动虽不能显著提升睾丸的重量,但可明显提升睾丸系数。贾斯媛[13]发现游泳运动会使肥胖雄性小鼠黄体生成素和卵泡刺激素显著增加,分析可能由于运动使肥胖雄性小鼠体内的GnRH得到良性刺激,进而促进其分泌黄体生成素和卵泡刺激素。我们先前的研究发现[3]经过中等负荷有氧运动后,肥胖运动组大鼠性激素水平有明显上升,精子质量显著提高,提示运动使大鼠的促性腺激素释放激素受到良性刺激,从而促进黄体生成素和卵泡刺激素的合成与释放,进而刺激睾酮的分泌,改善了促进精子发育的激素环境,提高了精子的质量。
3.2 有氧运动对大鼠生精细胞凋亡的影响及机制
3.2.1 肥胖对大鼠生精细胞凋亡的影响及其机制
已证实生殖细胞凋亡要借助两大途径:线粒体通路和死亡受体途径,而Caspase-3作为死亡受体途径中最后一个环节,在调控生殖细胞的凋亡上发挥了决定性的作用。目前,有关肥胖对Caspase-3的研究较少。郭银谋[15]发现肥胖人群的乳腺癌组织Caspase-3的蛋白水平明显高于正常体重组,且Caspase-3的水平与乳腺癌细胞凋亡之间存在正相关关系。本实验发现,与正常组相比较,肥胖组大鼠的睾丸组织Caspase-3的水平显著升高,精子数量明显下降,提示肥胖可能会通过上调Caspase-3水平而增加生精细胞的凋亡。
肿瘤坏死因子(TNF-α)可激活死亡受体途径,最终作用到Caspase-3,引发细胞凋亡。肿瘤坏死因子作为一种具有多种生物活性的细胞因子,除了脂肪组织是其主要来源外,近些年发现生殖细胞同样也分泌肿瘤坏死因子,如支持细胞、圆形精子细胞等。睾丸中的巨噬细胞、生精细胞能分泌大量TNF-α,可以影响睾丸间质细胞的睾酮生物合成过程,通过参与精子的发生与凋亡,对生殖功能产生影响。目前研究证实[16]适宜水平的TNF-α可以正向调节睾丸的功能,而含量过高则容易抑制睾酮的生成,明显降低睾丸生殖细胞的生存率,从而诱发其凋亡。有关肥胖对TNF-α的影响,多数认为啮齿类动物脂肪中TNF-α都过量表达,肥胖可以促进脂肪细胞表达释放TNF-α。有学者[16]对单纯性肥胖儿童的血液进行检测,发现TNF-α水平显著高于正常儿童。动物研究发现[7,15]与正常组大鼠相比较,肥胖组大鼠的TNF-α水平明显升高,睾丸组织形态学发生改变,可能与肥胖促进TNF-α含量上升有关。本实验研究也发现同正常组相比,肥胖大鼠的睾丸组织中TNF-α的基因表达水平显著上升,同时伴有睾酮水平下降和精子的数量、活力下降。
在哺乳动物的睾丸组织中,TNF-α促进生殖细胞程序性死亡,通过诱导死亡受体信号各级因子引发细胞凋亡。最典型的死亡受体是自杀相关因子(factor associated suicide,Fas,也称CD95或Apo1)。1989年Yonehara等发现了一株单克隆抗体,可以识别一种表达于髓样细胞、T淋巴细胞和成纤维细胞表面的未知分子,诱导多种人细胞系发生凋亡。这种新的膜分子被称为Fas。同年,Trauth等也发现了一株可以诱导活化或恶性变淋巴细胞凋亡的单克隆抗体,他们将这株抗体所识别的蛋白称为凋亡蛋白-1(Apo-1)。Fas属于肿瘤坏死因子家族,是一个细胞表面分子,在凋亡信号转导中起关键作用。
Fasl是Fas的天然受体,能够特异性地与靶细胞上的Fas结合诱导细胞凋亡。Fas主要在胸腺及睾丸组织有所表达。有报道[19-20]若Fasl/Fas系统异常升高,TNF-α可通过生殖细胞膜上的Fas与其受体结合,形成能传递信号的三聚体。三聚体通过募集死亡结构阈形成二聚体而发挥作用,凋亡作用明显增强,最终通过激活Caspase-3引起生殖细胞大量凋亡,导致精子减少或发育障碍,甚至男性不育。马丁[16]研究发现肥胖组大鼠睾丸组织中的Fas、Caspase-3水平显著上升,生殖细胞凋亡率显著升高。曹翔[19]等通过体外培育大鼠的内皮细胞,并对其凋亡情况进行测定,结果发现肿瘤坏死因子可以提高内皮细胞Fas的表达,在TNF-α作用下Fas表达加强,进一步诱导细胞凋亡。在免疫性睾丸炎中,TNF-α可以提高Fas表达,引发生精细胞凋亡。可见生殖细胞产生的TNF-α可通过调节Fas水平从而控制其凋亡。本研究发现,肥胖组大鼠的睾丸组织中FasmRNA转录水平显著升高,并且与TNF-α和Caspase-3 mRNA表达的增加相一致,推测肥胖可能会通过死亡受体途径造成生殖细胞大量凋亡。武进峰[20]等人对大鼠进行10周的高脂喂养,造模成功后,对实验组大鼠的生殖细胞经过RT-PCR和免疫组化测定后发现睾丸中Fas水平明显大于对照组。李奇[17]也对肥胖大鼠睾丸中的Fas进行检验,发现同对照组进行比较,FasmRNA含量显著上升。
3.2.2 有氧运动对肥胖大鼠生精细胞凋亡的影响及其机制
本研究发现Caspase-3在正常运动组和肥胖运动组大鼠的睾丸组织中基因转录水平显著低于对照组,同时精子的数量显著性增加,推测8周的跑台运动缓解了睾丸生殖功能障碍,下调Caspase-3的水平,降低精子细胞的凋亡率。有研究表明[4],对大鼠进行12周的脚踏车运动,发现Caspase-3及上游的几种促凋亡、抗凋亡蛋白水平发生了改变,凋亡程序中的关键因子Caspase-3下调到原来的95%。陈小燕[21]对高脂膳食大鼠施加游泳运动,发现与肥胖组相比,肥胖运动组的Caspase-3的基因水平显著下降,同时大鼠的精子凋亡率明显下降。推测8周的跑台运动缓解了睾丸生殖功能障碍的情况,下调Caspase-3的水平,降低了精子细胞的凋亡率。
TNF-α可以诱导死亡受体途径,最终作用到Caspase-3而影响精子细胞凋亡。有关运动对TNF-α影响的结果不尽一致。有研究表明连续高强度运动能够使血浆TNF-α水平显著提升。但有研究表明[1-2]持续锻炼会使肥胖个体TNF-α水平明显下降。陈小燕发现[21]10周渐增游泳运动的大鼠与普通安静组相比,血清中的TNF-α水平降低。毕业[16]和胡玉龙[9]的研究也得到了类此结果。但是目前对于肥胖状态下有氧运动对睾丸组织内的TNF-α的基因表达研究尚少。本研究结果显示正常运动组与肥胖运动组相比,睾丸组织中的TNF-α的mRNA基因表达较低;正常运动组TNF-α的mRNA转录水平要比正常组低;肥胖运动组的TNF-α的mRNA转录水平较肥胖组显著下降,精子质量显著提升。
大量研究显示[13-14]有氧运动可以降低睾丸组织中的TNF-α水平。但是有关运动对Fas转录水平的调节结果还不一致。有报道证实[10]一次性力竭运动能够使大鼠FasmRNA水平增加,从而使生殖细胞凋亡率显著上升。李奇[17]对肥胖大鼠施加有氧运动干预后,发现睾丸中Fas水平明显下调的同时,终末剪切酶Caspase-3含量也有所降低,检验后发现睾丸组织中生精细胞凋亡率显著下降。Caspase-3水平的降低减轻了其对生殖细胞凋亡的促发作用。本研究结果显示与正常组和肥胖组相比,正常运动组和肥胖运动组睾丸TNF-α、Fas、Caspase-3 mRNA水平都明显下降,精子数量增加,改善了睾丸生殖细胞大量凋亡的现象。提示有氧运动可以降低死亡受体途径各级因子水平,减少对细胞凋亡最重要的终末剪切酶的促发作用,缓解了肥胖引起的生精细胞凋亡和雄性生殖功能障碍。
4 结论
1)饮食性肥胖可通过上调睾丸TNF-αmRNA表达,启动死亡受体途径的信号转导,抑制睾丸的发育,对雄性生殖功能造成负面影响。
2)长期有氧运动可降低TNF-αmRNA表达,抑制死亡受体途径的信号转导,改善肥胖雄性大鼠对睾丸功能的负面影响,提升了正常雄性大鼠的生殖功能。
[1]J.Samavat,I.Natali,S.Degl’Innocenti,et al.Acrosome reaction is impaired in spermatozoa of obesemen:a preliminary study[J].Fertility and sterility,2014,102:1274-1281.
[2]F.Hammiche,J.S.Laven,J.M.Twigt,et al.Body mass index and central adiposity are associated with sperm quality inmen of subfertile couples[J].Human reproduction,2012,27:2365-2372.
[3]常波,赵大林,康媛,等.8周跑台运动干预对雄性肥胖大鼠生殖功能的影响[J].广州体育学院学报,2016(6):90-93.
[4]Villance.Voluntary wheel running decreases Adiponectinse tissue mass and expression of leption mRNA in casbome-mendelrats[J].lipid,2014(49):85-96.
[5]BE Levin,AA Dunn-Meynel.Differential effects of exercise on body weight gain and adiposity in obesity proneand resistantrats[J].International Journal of Obesity,2006(30):722-727.
[6]马智,王富兰,胡敏.体质量指数腰围及腰臀比对精子的影响[J].现代医药卫生,2014,30(13):1927-1928,1932.
[7]FuiMN,Dupuis P,Grossmann M.Lo were d testosterone inmale obesity:mechanisms,morbidity and management[J].Asian J Androl,2014(16):223-231.
[8]Kort H I,Massey JB,ElsnerCW,et al.Impact of body mass index values on sperm quantity and quality[J].JAndro,2006,27(3):450-452.
[9]胡玉龙,李宁川,李奇,等.有氧运动及消脂散对高脂膳食诱导的大鼠睾丸组织形态结构改变干预作用的研究[J].实用临床医药杂志,2012,16(22):5-8,13.
[10]李玲,李宁川,陆伟,等.有氧运动对高脂膳食诱导雄性大鼠生殖机能的干预作用[J].实用临床医用杂志,2014,18(11):10-13.
[11]徐庆阳,方翔,许苑,等.营养性肥胖对大鼠精子的影响[J].皖南医学院学报,2007,26(4):254-256.
[12]翟玲玲,赵剑,白英龙,等.肥胖对小鼠生殖功能影响及与性激素关系[J].中国公共卫生,2011,27(2):204-205.
[13]贾斯媛.不同负荷运动对肥胖雄性小鼠下丘脑脂联素-吻素-GnRH的影响[D].沈阳:沈阳体育学院,2015.
[14]田智谋.肥胖对大鼠生殖细胞线粒体凋亡途径的影响[D].太原:山西医科大学,2011.
[15]郭银谋,高建芝.Caspase-3在肥胖患者乳腺癌组织中的表达与细胞凋亡的关系[J].临床辅助检查,2011,32(13):233-234.
[16]毕业,李建设.有氧运动对单纯性肥胖儿童血中瘦素、神经肽、胰岛素和肿瘤坏死因子的影响研究[J].体育科学,2005,25(12):37-39.
[17]李奇.有氧运动联合消脂散对高脂膳食雄性大鼠生殖机能的干预作用[D].扬州:扬州大学,2012.
[18]马丁.营养性肥胖对青春期大鼠睾丸生殖细胞中Fas/FasL凋亡途径的影响[D].太原:山西医科大学,2012.
[19]曹翔,金惠铭.Fas、NF-κB及Caspases在TNFα介导的微血管内皮细胞凋亡中的作用[J].中国病理生理杂志,2001,17(8):15-18.
[20]武进峰,马丁,李承勇,等.营养性肥胖对大鼠睾丸生殖细胞Fas/FasL基因表达及凋亡的影响[J].中国药物与临床,2011,11(4):404-406.
[21]陈小燕.有氧运动对雄性高脂膳食大鼠生殖细胞凋亡的研究[D].扬州:扬州大学,2014.
