百里醌通过磷酸化p38MAPK途径介导NSCLC细胞毒性作用的机制研究
2018-06-19陈子盛廖小雯张亦飞肖靖华刘清霞车鹏彪朱连雨田东波
陈子盛,廖小雯,张亦飞,肖靖华,陈 芸,刘清霞,王 鹏,车鹏彪,朱连雨,田东波
(1.广州医科大学附属第六医院/清远市人民医院 呼吸二病区,广东 清远,511500; 2.广东省五邑中医院 神经内科,广东 江门,529000)
肺癌是目前全球最常见的恶性肿瘤之一,研究[1]表明2012年全球新发肺癌患者1.82/百万,死亡1.59/百万,肺癌约占所有肿瘤相关性死亡的18%。非小细胞肺癌(NSCLC)占所有肺癌的80%~85%,Ⅰ期和Ⅱ期NSCLC单纯根治术后5年存活率约70%,而晚期NSCLC 5年存活率<5%[2]。目前中国晚期NSCLC除针对驱动基因精准靶向治疗,最常用一线化疗是含铂两药方案,即长春瑞滨、吉西他滨、多西他赛、紫杉醇、替吉奥联合铂类,复发转移、治疗失败的NSCLC,通常是对以铂类为基础的化疗方案不应答或耐药[3],因而急需寻找新治疗方法,以降低一线方案耐药或提供有效稳定新靶点。
百里醌(TQ))是黑种草籽(Nigella damascena)挥发油的主要生物活性成分,有研究[4-5]表明TQ具有抗炎、抗氧化、抗高血压、抗菌、抗哮喘、抗肿瘤等功效,TQ能抑制乳腺癌、白血病、黑色素瘤、结肠癌、宫颈癌、多耐药卵巢癌、前列腺癌细胞增殖,且减少顺铂化疗方案不良反应如心肌损伤[6],并对正常细胞具有较高剂量耐受性[7],提示TQ具有较好安全性,有望成为NSCLC治疗选择。本研究探讨TQ对NSCLC毒性作用的分子机制,现报告如下。
1 材料与方法
1.1 试剂与细胞培养
SK-MES-1、95-D肺鳞癌细胞和A549肺腺癌细胞,均购自中国科学院细胞库(上海),TQ(纯度≥98%)、U0126和SB203580均购自Sigma,实验所需抗体p-p38、p38,p-ERK1/2、ERK1/2,p-JNK、JNK,β-actin均购自Cell Signaling Technology,化学发光试剂盒购自Millipore。SK-MES-1、95-D、A549培养条件均为: DMEM培养基(高糖,含10%胎牛血清,不加双抗),5%CO2,37.0 ℃。
1.2 实验设计及分组
采用DMSO溶解TQ后配成100 mmol/L,按梯度稀释法配成工作液浓度,对照组加入同体积的DMSO。
1.3 MTT测细胞毒性
SK-MES-1、95-D、A549细胞接种于96孔板,5 000个/孔,细胞贴壁后同步化12 h,分别加20、40、60、80、100 μmol/L的TQ,每个浓度设6复孔,孵育24 h后吸掉培养基,每孔加PBS稀释的1×MTT液100 μL培养4 h,弃上清,每孔加100 μL DMSO,震荡使结晶充分溶解,酶标仪570 nm测吸光度OD值,细胞存活率%=OD实验组/OD对照组×100%,利用excel回归方程计算TQ的IC50值。SK-MES-1时间依赖性实验细胞接种密度同上,予约TQ IC50浓度分别作用10 min、24 h、48 h,MTT检测方法如上计算细胞存活率。
1.4 Western blot实验
将指数生长期的SK-MES-1细胞接种于10 cm2培养皿,密度为1.5×106个/皿,贴壁后同步化12~16 h,处理组予10 μmol/L的U0126预处理1 h后加入TQ孵育30 min,对照组予DMSO处理,提取总蛋白,采用BCA法测样品总蛋白浓度,按Western blot实验方法进行电泳、转膜、5%脱脂牛奶封膜,一抗为羊抗兔p-p38、p38,p-ERK1/2、ERK1/2,p-JNK、JNK,4℃孵育过夜或者室温1 h,TBST清洗后室温下兔二抗孵育30 min,最后利用化学发光试剂盒显影并采集图像。
1.5 统计学分析
2 结 果
2.1 不同浓度TQ对NSCLC细胞毒性作用
分别予20、40、60、80、100 μmol/L TQ孵育SK-MES-1细胞24 h,可见TQ呈浓度依赖性发挥细胞毒性作用,通过Excel回归方程式计算,TQ IC50≈66 μmol/L,予60 μmol/L TQ分别作用10 min、24 h、48 h,可见TQ呈时间依赖性发挥细胞毒性作用(图1A、B),扩展至95-D和A549细胞,同样观察到TQ呈浓度依赖性降低细胞存活率(图1C、D)。

A.TQ呈浓度依赖性发挥SK-MES-1细胞毒性作用,***P=0.000;

B.TQ呈时间依赖性发挥SK-MES-1细胞毒性作用,***P=0.000;
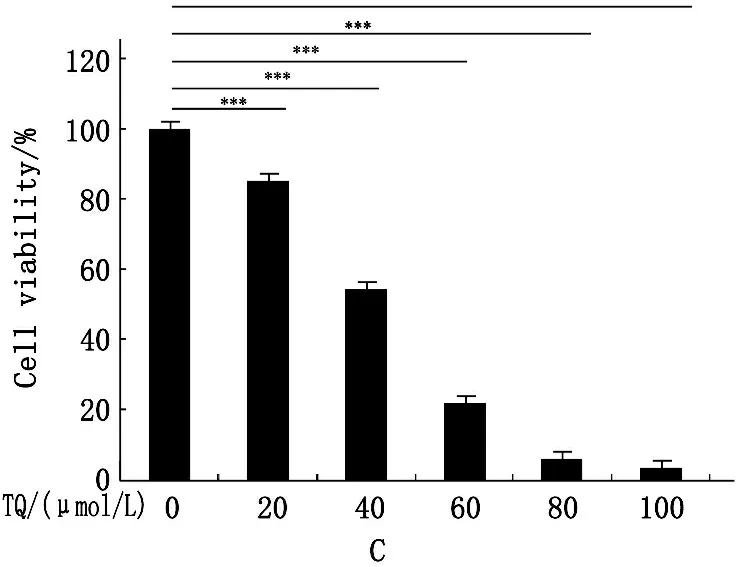
C.TQ呈浓度依赖性发挥95-D细胞毒性作用,***P=0.000;
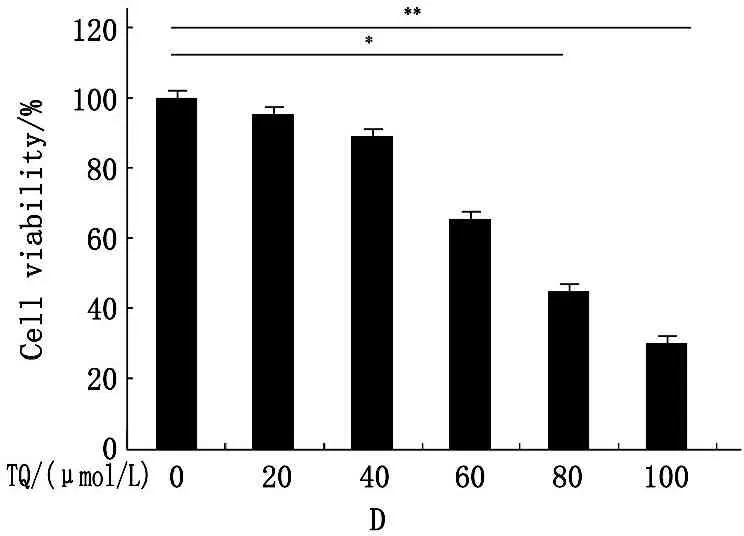
D.TQ呈浓度依赖性发挥A549细胞毒性作用,*P<0.05,**P<0.01。
图1不同浓度TQ对NSCLC的细胞毒性作用
2.2 TQ通过p38途径介导NSCLC细胞毒性作用
为验证TQ是否通过p38途径介导NSCLC细胞毒性作用,对照组予30和60 μmol/L TQ作用于SK-MES-1 1h; 实验组分别予10 μmol/L U0126和10 μmol/L SB203580预处理1 h后,分别加入30和60 μmol/L TQ 1 h,吸掉含药培养基,加正常培养基,培养24 h后行MTT检测。结果表明,30 μmol/L TQ和10 μmol/L U0126+30 μmol/L TQ比较其细胞存活率有显著差异(P=0.000),但与10 μmol/L SB203580+30 μmol/L TQ比较无差异(P=1.00),10 μmol/L SB203580+30 μmol/L TQ与10 μmol/L U0126+30 μmol/L TQ组比较有显著差异(P=0.000); 随着TQ浓度增加,60 μmol/L TQ、10 μmol/L U0126+60 μmol/L TQ、10 μmol/L SB203580+60 μmol/L TQ组两两比较,其细胞存活率均无显著差异(P>0.05),表明即使低剂量TQ状态下,ERK抑制剂U0126也不能阻断TQ的细胞毒性作用,即TQ不是通过活化ERK1/2起效,但p38抑制剂SB203580则明显降低TQ的细胞毒性作用,表明TQ可快速(1 h)通过p38途径介导细胞毒性作用,可是随TQ浓度升高接近IC50值,SB203580短暂抑制不能明显保护SK-MES-1(图2)。
予10 μmol/L SB203580预处理95-D细胞1 h后,分别加入按梯度稀释的5~40 μmol/L TQ孵育1 h,换正常培养基培养24 h。10 μmol/L SB203580+40 μmol/L TQ组为10 μmol/L SB203580预处理1 h,加40 μmol/L TQ(≈95-D的TQ IC50值)孵育1 h后,更换含10 μmol/L SB203580的正常培养基培养24 h。MTT结果表明,随TQ浓度增加,SB203580对95-D细胞保护作用逐渐减弱,10 μmol/L SB203580+40 μmol/L TQ组与40 μmol/L TQ组比较其细胞存活率仍有显著差异(P=0.033),表明持续p38抑制仍能明显降低TQ对95-D细胞毒性作用(图3)。

∗P=0.016, ∗∗∗P=0.000。图2 TQ透过p38介导SK-MES-1细胞毒性作用

∗P=0.033。图3 TQ通过p38途径介导95-D细胞毒性作用
2.3 TQ对NSCLC细胞毒性作用的分子机制
Western blot结果表明U0126能显著抑制ERK1/2磷酸化,p38随TQ浓度增加而磷酸化增加,但ERK1/2磷酸化减少,JNK磷酸化无明显变化,表明TQ通过磷酸化p38介导细胞毒性作用,而不是ERK1/2(图4)。

图4 TQ对NSCLC细胞毒性作用的分子机制
3 讨 论
丝裂原活化蛋白激酶(MAPK)是一组丝氨酸-苏氨酸蛋白激酶,刺激信号如生长因子、细胞因子、射线、基因毒性化疗药、应激、渗透压等均可激活MAP3K,产生级联效应,活化MAPK(ERK、JNK/SAPK、p38),MAPK一旦被胞外刺激信号激活将广泛调节细胞增殖、迁移、凋亡、分化和衰老[8]。许多化疗药需激活p38才能发挥作用,通过重新激活p38诱导癌细胞凋亡被认为是治疗NSCLC的重要途径之一[9],p38活化增强化疗敏感性,促进癌细胞凋亡,引起癌细胞周期停滞,抑制癌细胞生长和分化[10],比如在乳腺癌细胞中环磷酰胺、结肠癌细胞中奥沙利铂均通过活化p38通路而诱导癌细胞凋亡[11-12],依托泊苷增加NSCLC细胞MKK3/6-p38的磷酸化[13],活化的p38通过灭活Met受体增强顺铂化疗敏感性[14]。
TQ具有抗炎、抗氧化、抗肿瘤等功效,本研究表明TQ呈浓度及时间依赖性发挥NSCLC细胞毒性作用。为进一步验证TQ是否通过p38途径介导NSCLC细胞毒性效应,予10 μmol/L U0126和10 μmol/L SB203580预处理SK-MES-1,抑制ERK1/2和p38磷酸化,加入30 μmol/L TQ后孵育1 h后更换正常培养基培养24 h,结果表明U0126并不能防止TQ对SK-MES-1的毒性作用,但SB203580却对SK-MES-1有保护作用,其细胞存活率与正常组比较无显著差异,表明TQ可能通过磷酸化p38介导细胞毒性作用,但当TQ浓度接近IC50值时(60 μmol/L),10 μmol/L SB203580+60 μmol/L TQ组与10 μmol/L U0126+60 μmol/L TQ、60 μmol/L TQ组细胞存活率比较均无显著差异,表明当TQ达到一定浓度时,SB203580短时间抑制p38不再能阻断TQ毒性作用。对95-D予接近IC50值的TQ浓度40 μmol/L作用1 h后,更换含10 μmol/L SB203580的培养基培养24 h,与40 μmol/L TQ组比较,细胞存活率显著增加,表明持续p38抑制明显降低TQ对NSCLC细胞毒性作用。
进一步探讨TQ对NSCLC细胞毒性作用的分子机制,结果表明p38随TQ浓度增加而磷酸化增加,但ERK1/2磷酸化减少,而JNK磷酸化无明显改变,表明TQ透过磷酸化p38介导细胞毒性作用,同时抑制ERK1/2介导的细胞增殖信号,并非
如Yang J等[15]表明TQ通过磷酸化ERK1/2抑制NSCLC细胞增殖、迁移、侵袭。
综上所述,NSCLC细胞基本不表达p-p38,一旦其p38磷酸化,即可诱发NSCLC细胞毒性级联反应,TQ在短时间内(1 h)就能决定24 h后NSCLC细胞的存亡,同时抑制ERK1/2介导的细胞增殖信号。由此可见,TQ是一种极有前景、值得继续深入探讨的备选抗癌药或者辅助化疗增敏药。
[1] Ferlay J,Soerjomataram I,Dikshit R,et al.Cancer incidence and mortality worldwide: sources,methods and major patterns in GLOBOCAN 2012[J].Int J Cancer,2015,136(5): E359-386.
[2] Scrima M,Zito Marino F,Oliveira D M,et al.Aberrant Signaling through the HER2-ERK1/2 Pathway is Predictive of Reduced Disease-Free and Overall Survival in Early Stage Non-Small Cell Lung Cancer (NSCLC) Patients[J].Journal of Cancer,2017,8(2): 227-239.
[3] Ravdin P M,Davis G.Prognosis of patients with resected non-small cell lung cancer: impact of clinical and pathologic variables[J].Lung Cancer,2006,52(2): 207-212.
[4] Dastjerdi M N,Mehdiabady E M,Iranpour F G,et al.Effect of Thymoquinone on P53 Gene Expression and Consequence Apoptosis in Breast Cancer Cell Line[J].International journal of preventive medicine,2016,7: 66-74.
[5] Effenberger-Neidnicht K,Schobert R.Combinatorial effects of thymoquinone on the anti-cancer activity of doxorubicin[J].Cancer chemotherapy and pharmacology,2011,67(4): 867-874.
[6] Adali F,Gonul Y,Kocak A,et al.Effects of thymoquinone against cisplatin-induced cardiac injury in rats[J].Acta cirurgica brasileira,2016,31(4): 271-277.
[7] Abdelfadil E,Cheng Y H,Bau D T,et al.Thymoquinone induces apoptosis in oral cancer cells through p38beta inhibition[J].The American journal of Chinese medicine,2013,41(3): 683-696.
[8] Sui X,Kong N,Ye L,et al.p38 and JNK MAPK pathways control the balance of apoptosis and autophagy in response to chemotherapeutic agents[J].Cancer Lett,2014,344(2): 174-179.
[9] Olson J M,Hallahan A R.p38 MAP kinase: a convergence point in cancer therapy[J].Trends Mol Med,2004,10(3): 125-129.
[10] Cuadrado A,Nebreda A R.Mechanisms and functions of p38 MAPK signalling[J].Biochem J,2010,429(3): 403-417.
[11] Pang H,Cai L,Yang Y,et al.Knockdown of osteopontin chemosensitizes MDA-MB-231 cells to cyclophosphamide by enhancing apoptosis through activating p38 MAPK pathway[J].Cancer Biother Radiopharm,2011,26(2): 165-173.
[12] Chiu S J,Chao J I,Lee Y J,et al.Regulation of gamma-H2AX and securin contribute to apoptosis by oxaliplatin via a p38 mitogen-activated protein kinase-dependent pathway in human colorectal cancer cells[J].Toxicol Lett,2008,179(2): 63-70.
[13] Tsai M S,Weng S H,Chen H J,et al.Inhibition of p38 MAPK-dependent excision repair cross-complementing 1 expression decreases the DNA repair capacity to sensitize lung cancer cells to etoposide[J].Mol Cancer Ther,2012,11(3): 561-571.
[14] Lou X,Zhou Q,Yin Y,et al.Inhibition of the met receptor tyrosine kinase signaling enhances the chemosensitivity of glioma cell lines to CDDP through activation of p38 MAPK pathway[J].Mol Cancer Ther,2009,8(5): 1126-1136.
[15] Yang J,Kuang X R,Lv P T,et al.Thymoquinone inhibits proliferation and invasion of human nonsmall-cell lung cancer cells via ERK pathway[J].Tumour biology: the journal of the International Society for Oncodevelopmental Biology and Medicine,2015,36(1): 259-269.
