FOXG1在结直肠癌侵袭及转移中的作用及机制
2018-06-11吴海霞钱程刘春刚向俊宇叶迪张振芳张献全
吴海霞,钱程,刘春刚,向俊宇,叶迪,张振芳,张献全
1 重庆医科大学 附属第二医院肿瘤科,重庆 400010
2 第三军医大学 西南医院 生物治疗中心,重庆 400038
结直肠癌是全球范围内常见的恶性消化道肿瘤之一,其发病率和死亡率还在逐年上升[1]。目前结直肠癌的治疗措施主要为手术切除,但由于结直肠癌细胞具有较高的侵袭和转移的生物学特性,故影响结直肠癌患者治疗效果的主要因素为远处转移,以肝转移最为常见[2-3]。因此找寻新的肿瘤标志物及治疗靶标对早期诊断和及时制止其侵袭转移,对患者的预后具有至关重要的意义。
上皮-间质转化 (Epithelial-mesenchymal transition EMT) 是一种常见的病理生理学现象,是指上皮样细胞获得间质细胞特性的过程,发生EMT时,细胞结构改变、极性消失、细胞间连接减少而运动能力则增强,表现出类似间质细胞的表型和迁移能力,如上皮样标志物 E-cadherin表达降低,而间质样标志如Vimentin、Fibronectin则增高[4-8],近年来EMT被认为是肿瘤转移的起始环节,成为肿瘤转移研究的热点[9-11],因此对EMT的研究有助于增进对肠癌转移的认识。
叉头框G1 (Forkhead box G1,FOXG1) 基因,又名脑因子 1基因,是叉头框 (Forkhead box,FOX) 基因家族里的重要一员,位于14q12,其编码的转录因子常特异表达于人类脑组织,常与神经系统的发育性病变有关[12-17],有文献报道FOXG1在卵巢癌、肝癌中高表达,并可作为不良预后的分子指标[18-19],即 FOXG1通过抑制p21WAF1/CIP1转录,进而增强TGF-b介导的促肿瘤增殖能力抗增殖反应,但 FOXG1在结直肠癌中尚无报道,且 FOXG1是否能促进肿瘤的侵袭转移也未见报道。本研究首次分析了 FOXG1在结直肠癌侵袭转移中的作用及机制。
1 材料与方法
1.1 材料
各类肠癌细胞由本实验室保存,DMEM培养基、胎牛血清、胰酶、Lipofectamine 2000转染试剂等均购自 Gibco公司;兔抗人 FOXG1抗体购自 Abcam公司;兔抗人 E-cadherin、Vimentin、鼠抗人Fibronectin抗体购自Cell Signaling 公司;分子克隆和慢病毒包装等试剂由本实验室保存;Trizol试剂购自Invitrogen公司;逆转录聚合酶链式反应试剂盒购自 TaKaRa公司;所有引物均由金斯瑞公司负责合成。
1.2 方法
1.2.1 细胞培养
人结肠癌细胞系 RKO、SW480、SW620、LOVO、DLD-1培养于含10%胎牛血清1640培养基中 (含青链霉素) 在37 ℃、5% CO2的培养箱中孵育,当细胞密度达80%左右进行传代,本实验采用对数生长期中的细胞进行实验。
1.2.2 Western blotting检测
提取总蛋白,制10%胶进行SDS-PAGE,转膜2 h,5%脱脂牛奶封闭1 h,一抗孵育4 ℃过夜,一抗比例:GAPDH (1∶1 000)、FOXG1 (1∶700)、E-cadherin (1∶1 000)、Vimentin (1∶1 000)Fibronectin (1∶1 000),PBST洗1 h每10 min换一次液,二抗室温孵育1.5 h,PBST再洗1 h,拍照存图。
1.2.3 质粒的构建及慢病毒转染RKO细胞
设计并合成 3对 FOXG1的 shRNA片段(shFOXG1),运用DNA重组技术获得重组质粒,经双酶切技术及测序方法鉴定正确后进行慢病毒的包装,包装后存放于-80 ℃,细胞分为加入病毒的PLKO.1-shFOXG1实验组及未经处理的阴性对照组。转染前将细胞接种于6孔板中,当细胞密度达 50%左右进行细胞稳定转染,转染前 1 h常规换液,经嘌呤霉素筛选约1个月后可获得稳定的细胞株。
1.2.4RT-PCR检测
以Trizol法常规提取细胞中的总RNA,按试剂盒进行反转录反应合成cDNA,再以cDNA为模板进行实时荧光定量 PCR反应,反应条件为95 ℃ 60 s;95 ℃ 15 s,60 ℃ 60 s共 40 个循环,以 GAPDH作为内参,Bio-Rad 凝胶成像仪采集图片,以Folds=2-ΔΔCt来表示实验组和对照组中目的基因的表达倍数比[20],各引物序列见表1。
1.2.5 划痕实验
将细胞接种于6孔板中,待细胞融合率达到70%−80%时,使用枪头比着直尺用力划线,用PBS轻轻冲洗划下的细胞,加无血清培养基继续培养,分别在0 h和48 h拍照存图。
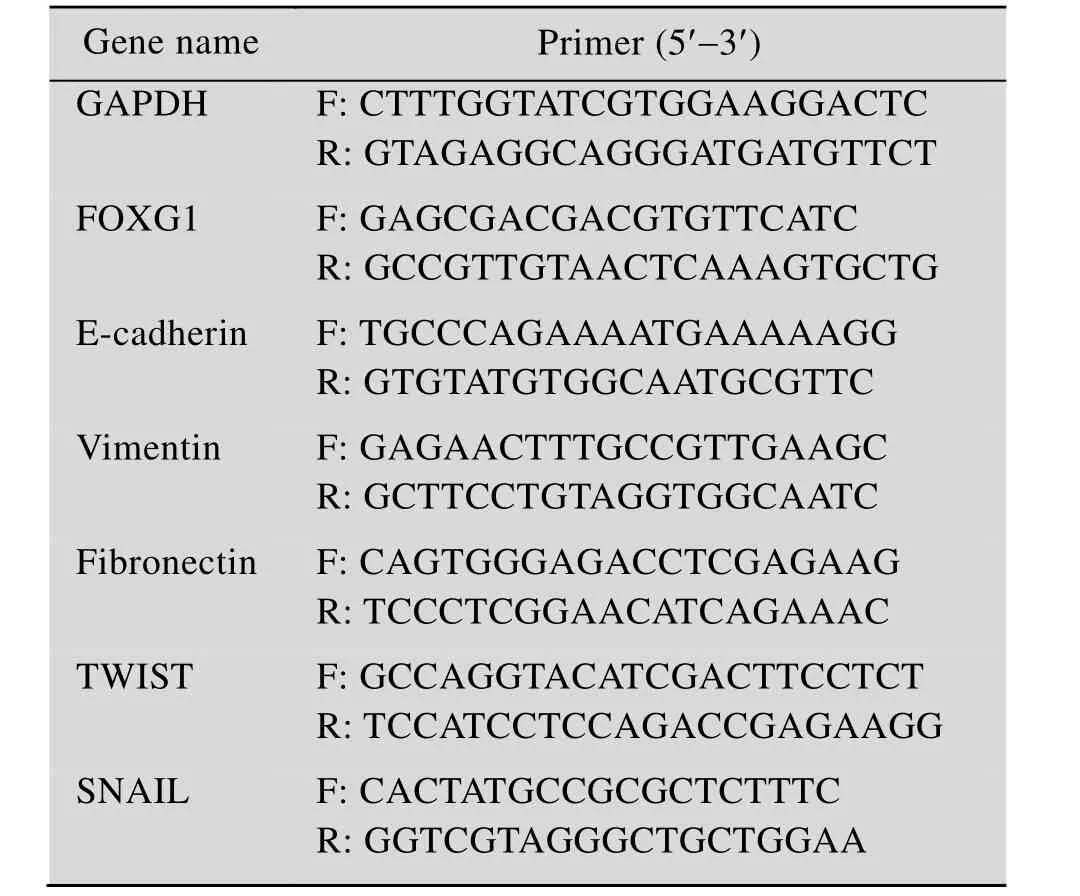
表1 目的基因的引物序列Table 1 The primers of related genes
1.2.6 Transwell检测
无血清培养基与Matrigel胶按1∶1配成基质胶,按10 μL/室加入包被Transwell小室底部膜的上室面,室温风干,上室面加入含10 000个细胞的细胞悬液,下室面加 900 μL的含 20%的 FBS的培养基,置于37 ℃、5% CO2的培养箱中孵育48 h后用4%多聚甲醛进行固定30 min,PBS洗涤2次,用结晶紫燃料染色30 min,轻擦上室面细胞,显微镜下计数穿过上室面得细胞数,随机取5个视野拍照取均值。细胞迁移试验:用未被包被的Transwell小室,转移时间为24 h,所铺细胞量为含5 000个细胞的细胞悬液。其余步骤同侵袭实验。
1.2.7 统计学方法
计量资料用±s表示,采用SPSS 13.0进行两独立样本的t检验,P≤0.05有统计学意义。
2 结果与分析
2.1 FOXG1在5种肠癌细胞系中的表达及与侵袭能力的相关性分析
在 RKO、SW480、SW620、LOVO、DLD-1五种肠癌细胞系中,通过Western blotting检测发现,FOXG1在RKO细胞中蛋白表达量最高,而在DLD-1细胞中表达最低 (图1A),由文献得知这 5种结直肠癌细胞的侵袭力如下:LOVO>SW620>RKO>SW480>DLD-1[21],通过对FOXG1蛋白相对表达量与相应细胞侵袭能力作相关性分析发现,在肠癌细胞中 FOXG1的表达量与侵袭能力呈正相关 (图1B)。
2.2 敲低FOXG1可抑制RKO细胞侵袭和迁移能力
重组质粒构建成功,经双酶切技术及测序方法鉴定为正确后进行慢病毒的包装、纯化及稳定转染,经筛选后获得了稳定的结直肠癌细胞株,再通过3种含shFOXG1沉默序列的慢病毒分别感染RKO细胞后,Western blotting检测发现,与对照组相比,shFOXG1-3干扰片段对FOXG1干扰沉默效果最明显 (图 2A)。划痕实验结果表明在RKO细胞中干扰FOXG1后,其迁移能力显著变弱(图2B)。Transwell结果表明干扰FOXG1后,其侵袭和迁移能力也显著下降 (图 2C),由于shFOXG1-3干扰片段对FOXG1干扰沉默效果最明显,故后续实验均以shFOXG1-3慢病毒转染细胞,并命名为干扰组 (Interfered group),单独使用时简写为shFOXG1。
2.3 过表达FOXG1促进DLD-1细胞侵袭和迁移能力
在DLD-1细胞过表达FOXG1后,与对照组相比较,FOXG1蛋白显著升高 (图3A)。RT-qPCR进一步验证过表达效率 (图 3B)。Transwell结果表明,过表达FOXG1促进了DLD-1细胞的侵袭和迁移能力 (图3C)。

图 1 在结直肠癌细胞中 FOXG1的表达与侵袭能力呈正相关Fig. 1 FOXG1 expression of colorectal cancer cells was positively correlated with their invasion ability. (A)FOXG1 expression in colorectal cancer cell lines were analyzed by Western blotting. (B) The correlation between FOXG1 expression and cell invasive abilities was analyzed (Pearson correlation assy).
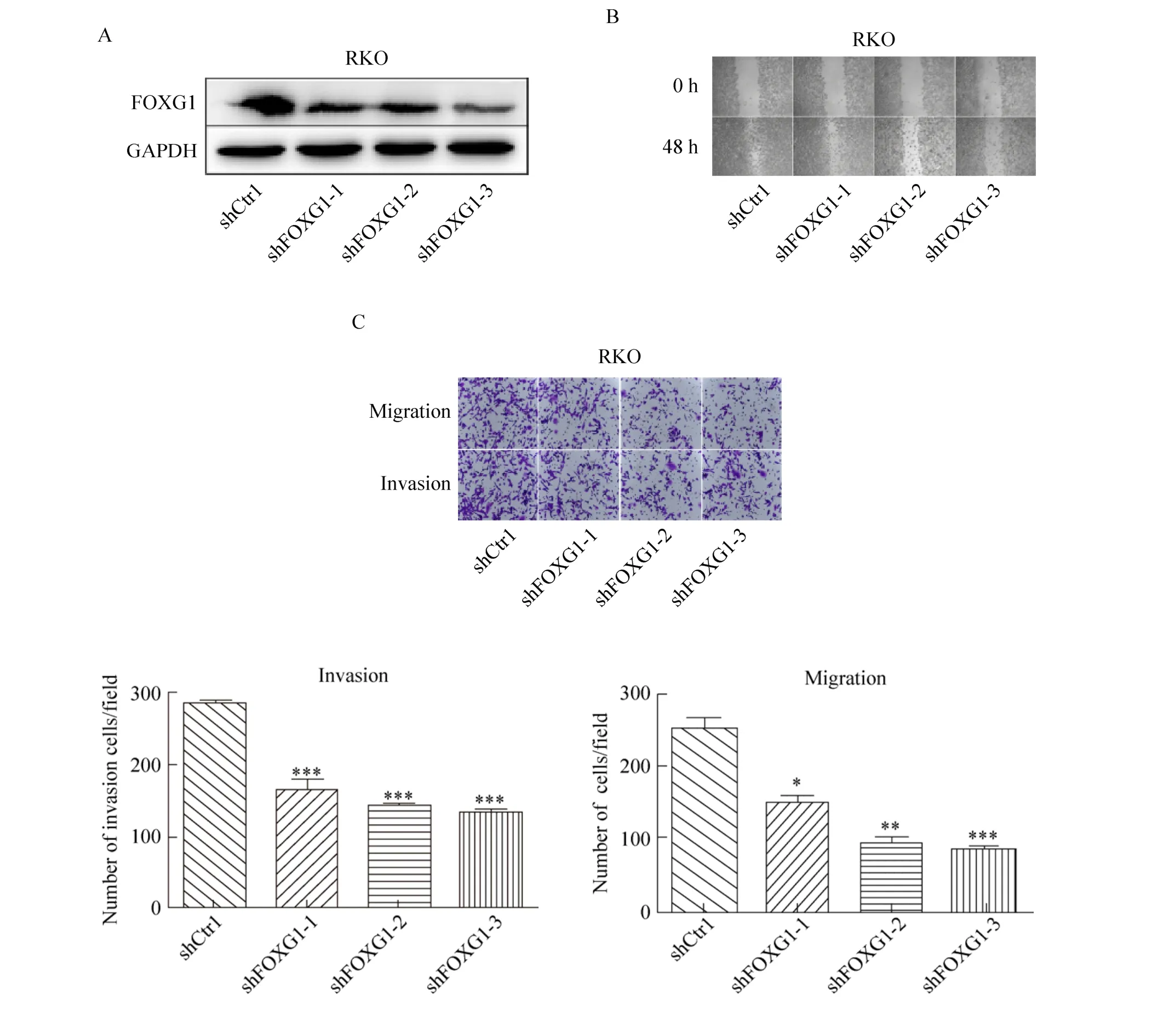
图2 干扰FOXG1抑制RKO细胞侵袭和迁移能力Fig. 2 Knockdown of FOXG1 weakened invasion and migration abilities of RKO cells. (A) The interference efficiency of FOXG1 protein was detected by Western blotting after three virus-infected RKO cells containing shFOXG1 silencing sequence (P<0.05). (B) Healing ability was tested by scratch test. (C) Transwell assay show invasion and migration abilities of FOXG1 Knockdown RKO cells compared with corresponding control cells. *: P<0.05;**: P<0.01; ***: P<0.001.
2.4 FOXG1对维持结肠癌EMT具有重要作用
干扰 FOXG1后细胞形态由典型的长梭型变成类圆形或多边形,生长方式也由较为分散转为较密集的生长 (图 4A)。干扰 FOXG1,Western blotting检测分析发现上皮样标志物E-cadherin表达在mRNA和蛋白水平都增加,而间质类标志物Vimentin、Fibronectin表达降低 (图4B)。而过表达FOXG1则相反 (图4C)。
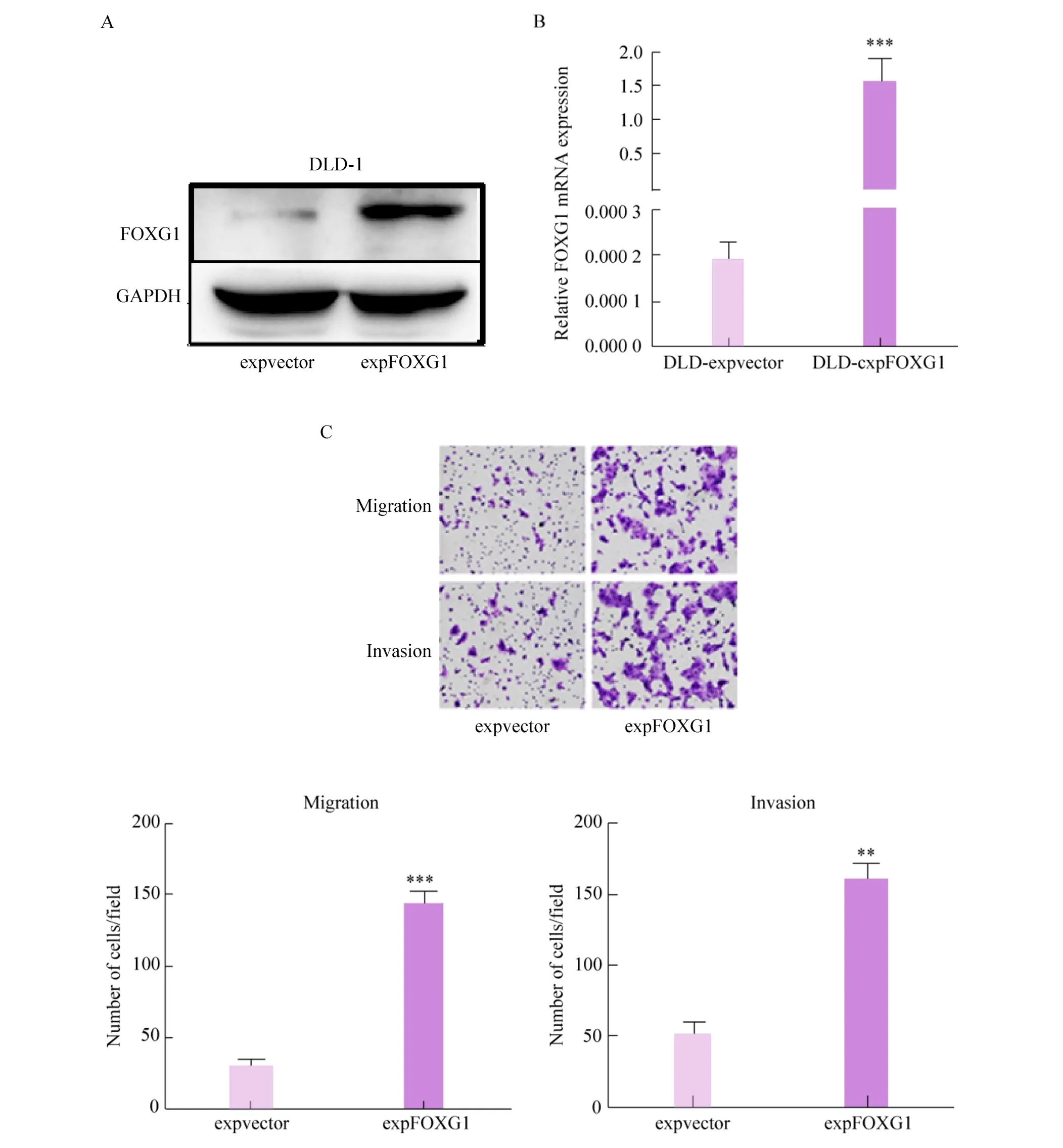
图3 过表达FOXG1促进DLD-1细胞侵袭和迁移能力Fig. 3 Overexpression of FOXG1 promotes invasion and migration abilities of DLD-1 cells. (A) The overexpression efficiency of FOXG1 protein was detected by Western blotting. (B) The overexpression efficiency of FOXG1 mRNA was detected by RT-qPCR. (C) Transwell assay show invasion and migration abilities of FOXG1 overexpression DLD-1 cells compared with corresponding control cells. **: P<0.01; **: P<0.001.

图4 FOXG1对维持EMT具有重要作用Fig. 4 FOXG1 plays an important role in maintaining EMT. (A) Morphological changes in RKO cells after stable FOXG1 expression deletion. (B) Expression changes of EMT markers were analyzed by RT-qPCR and Western blotting in RKO cells after FOXG1 knockdown. (C) Expression changes of EMT markers were analyzed by RT-qPCR and Western blotting in DLD-1 cells after overexpression of FOXG1. *: P<0.05; **: P<0.01; ***: P<0.001.
3 讨论
研究表明FOX基因家族与肿瘤EMT相关,如FOXQ1、FOXC1、FOXC2、FOXM1能够降低E-cadherin表达,促进肿瘤的转移并发生EMT[22-23]。FOXG1是 FOX基因家族里的重要一员,位于14q12,其编码的转录因子常特异表达于人类脑组织,常与神经系统的发育性病变有关[12],它涉及神经细胞的分化、增殖、转移和凋亡,对端脑的发育有重要作用[24],其异常表达常致 FOXG1的相关疾病[25-26],如发育迟缓、语言障碍、智力低下等。有研究表明,FOXG1在卵巢癌、肝癌中高表达并作为促癌基因,通过对P21的抑制促进癌细胞的增殖[18-19],这提示我们 FOXG1可能与肿瘤的发生发展有关。而 FOXG1在结直肠癌中表达情况尚无报道,且之前对于 FOXG1在某些实体瘤的研究主要局限于增殖方向,FOXG1是否能促进肿瘤的其他生物学功能还不明确,且大量研究表明 EMT与结直肠癌的侵袭和转移有关[27],故本研究在细胞水平上探索了在直结肠癌中FOXG1与EMT和转移的关系。基于上述,查阅TCGA数据库得知在结直肠癌组织中 FOXG1的表达高于正常组织,同时本研究检测了5种结直肠癌中 FOXG1的蛋白表达水平,并作了蛋白相对表达量与细胞侵袭能力相关性分析 (Pearson相关性检验),最终确定干扰和过表达的细胞系即对RKO细胞进行干扰,DLD-1细胞进行过表达,并检测对结直肠癌细胞侵袭转移能力的影响。结果表明慢病毒干扰载体PLKO.1-shFOXG1成功沉默了FOXG1的表达后,RKO细胞的侵袭转移能力下降,干扰FOXG1后RKO细胞形态由典型的长梭型变成类圆形或多边形,细胞形态变得饱满,细胞间的紧密连接增多生长也变得较为密集,且上皮类标志物 E-cadherin表达增高,而间质类标志物 Vimentin、Fibronectin表达降低,过表达组则相反,这些结果表明 FOXG1可能是肿瘤细胞维持间质样表型所必需的,但具体机制还需进一步研究。
综上所述,本研究证明 FOXG1能促进结直肠癌细胞侵袭和转移,并促进其发生EMT,有望成为一个新的治疗靶点,但其具体机制还需进一步探究。
[1]Kim JC, Kim SY, Roh SA, et al. Gene expression profiling: canonical molecular changes and clinicopathological features in sporadic colorectal cancers. World J Gastroenterol, 2008, 14(43):6662–6672.
[2]Sahani DV, Bajwa MA, Andrabi Y, et al. Current status of imaging and emerging techniques to evaluate liver metastases from colorectal carcinoma. Ann Surg, 2014,259(5): 861–872.
[3]Zhou ZW, Ren JQ, Wan DS, et al. Multivariate regressive analysis of prognosis of liver metastases from colorectal cancer. Chin J Cancer, 2006, 25(9):1149–1152 (in Chinese).周志伟, 任镜清, 万德森, 等. 结直肠癌肝转移患者预后的多因素分析. 癌症, 2006, 25(9): 1149–1152.
[4]Tiwari N, Gheldof A, Tatari M, et al. EMT as the ultimate survival mechanism of cancer cells. Semin Cancer Biol, 2012, 22(3): 194–207.
[5]Ahmed S, Nawshad A. Complexity in interpretation of embryonic epithelial-mesenchymal transition in response to transforming growth factor-β signaling.Cells Tissues Organs, 2007, 185(1/3): 131–145.
[6]Zhang JP, Zeng C, Xu L,et al.MicroRNA-148a suppresses the epithelial-mesenchymal transition and metastasis of hepatoma cells by targeting Met/Snail signaling. Oncogene, 2014, 33(31): 4069–4076.
[7]Lamouille S, Xu J, Derynck R. Molecular mechanisms of epithelial-mesenchymal transition. Nat Rev Mol Cell Biol, 2014, 15(3): 178–196.
[8]Theys J, Jutten B, Habets R, et al. E-cadherin loss associated with EMT promotes radioresistance in human tumor cells. Radiother Oncol, 2011, 99(3):392–397.
[9]Peng M, Hu YK, Song W, et al.MIER3 suppresses colorectal cancer progression by down-regulating Sp1,inhibiting epithelial-mesenchymal transition.Sci Rep,2017, 7(1): 11000.
[10]Li H, Yu BQ, Li JF, et al. Overexpression of lnc RNA H19 enhances carcinogenesis and metastasis of gastric cancer. Oncotarget, 2014, 5(8): 2318–2329.
[11]Yang X.A new miRNA story about tumor metastasis.Chin J Biotech, 2012, 28(2): 131–133 (in Chinese).杨晓. 肿瘤转移的新 miRNA故事. 生物工程学报,2012, 28(2): 131–133.
[12]Brancaccio M, Pivetta C, Granzotto M, et al.Emx2andFoxg1inhibit gliogenesis and promote neuronogenesis.Stem Cells, 2010, 28(7): 1206–1218.
[13]Jacob FD, Ramaswamy V, Andersen J, et al. Atypical Rett syndrome with selective FOXG1 deletion detected by comparative genomic hybridization: case report and review of literature. Eur J Hum Genet, 2009, 17(12):1577–1581.
[14]Florian C, Bahi-Buisson N, Bienvenu T.FOXG1-related disorders: from clinical description to molecular genetics. Mol Syndromol, 2012, 2(3/4/5):153–163.
[15]Ellaway CJ, Ho G, Bettella E,et al. 14q12 microdeletions excluding FOXG1 give rise to a congenital variant Rett syndrome-like phenotype. Eur J Hum Genet, 2013, 21(5): 522–527.
[16]Seltzer LE, Ma M, Ahmed S, et al. Epilepsy and outcome in FOXG1-related disorders. Epilepsia, 2014,55(8): 1292–1300.
[17]Kumakura A, Takahashi S, Okajima K, et al. A haploinsufficiency of FOXG1 identified in a boy with congenital variant of Rett syndrome. Brain Dev, 2014,36(8): 725–729.
[18]Adesina AM, Nguyen Y, Guanaratne P, et al.FOXG1is overexpressed in hepatoblastoma. Hum Pathol, 2007,38(3): 400–409.
[19]Chan DW, Liu VWS, To RMY, et al. Overexpression of FOXG1 contributes to TGF-βresistance through inhibition of p21WAF1/CIP1expression in ovarian cancer.Br J Cancer, 2009, 101(8): 1433–1443.
[20]Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods, 2001, 25(4): 402–408.
[21]Su L, Luo YL, Yang Z, et al. MEF2D transduces microenvironment stimuli to ZEB1 to promote epithelial-mesenchymal transition and metastasis in colorectal cancer. Cancer Res, 2016, 76(17):5054–5067.
[22]Xia LM, Huang WJ, Tian DA, et al.Overexpression of forkhead box C1 promotes tumor metastasis and indicates poor prognosis in hepatocellular carcinoma.Hepatology, 2013, 57(2): 610–624.
[23]Qiao YY, Jiang X, Lee ST, et al. FOXQ1 regulates epithelial-mesenchymal transition in human cancers.Cancer Res, 2011, 71(8): 3076–3086.
[24]Manuel MN, Martynoga B, Molinek MD, et al. The transcription factor Foxg1 regulates telencephalic progenitor proliferation cell autonomously, in part by controlling Pax6 expression levels. Neural Dev, 2011, 6: 9.
[25]McMahon KQ, Papandreou A, Ma M, et al. Familial recurrences ofFOXG1-related disorder: evidence for mosaicism. Am J Med Genet A,2015, 167(12):3096–3102.
[26]Golso ML, Kaestner KH. Fox transcription factors:from development to disease. Development, 2016,143(24): 4558–4570.
[27]Iwatsuki M, Mimori K, Yokobori T, et al. Epithelialmesenchymal transition in cancer development and its clinical significance. Cancer Sci, 2010, 101(2):293–299.
