望谟红球油茶种实表型变异分析
2018-06-08,,
, ,
(贵州省林业科学研究院, 贵阳 550005)
贵州独特的地质地貌及气候类型,造就了油茶生境的多样性,长期的地理阻隔及对生境的适应,加之油茶为异花虫媒受粉树种,导致杂种后代的性状类型丰富,为选育优质、高产及高抗的优良种质提供了丰富的材料,育种潜力大。
望谟红球油茶是贵州省册亨、望谟地区分布的特殊油茶类型,丰产稳产性能突出,对当地夏季干热的特殊气候类型有较强的适应能力。册亨县是望谟红球油茶的主产区之一,望谟红球油茶是该地主要栽培的农家品种,具有果实大,果皮革质光滑红润,植株抗病虫能力强及早实丰产的优良特性[1-2]。20世纪70~80年代,由贵州省林业科学研究院选育并参加了全国区试,望谟红球油茶被评为全国优良农家品种。陈晓春[1]对望谟红球油茶进行评价,认为望谟红球油茶具有速生早实、丰产、抗病性强等优点;罗在柒等[2]根据望谟红球油茶品种特性,对经济性状和形态指标进行了矩阵关联分析,初步确定了望谟红球油茶优树的选择标准。然而对于望谟红球油茶种质资源表型多样性的研究颇为有限。
表型性状多样性是植物遗传多样性的一个重要组成部分,植物的表型性状包括叶、花、种实及树体性状等,但对于产果类经济树种来说,种实性状尤为重要[3]。望谟红球油茶生产的目的是获取茶籽,其种实性状直接决定着茶籽的质量,而果实形态特征是最直观、最易于辨别的质量性状。本研究结合多重比较、主成分分析、聚类分析等方法,综合分析望谟红球油茶种实性状的表型多样性,为望谟红球油茶种质资源的保存利用和遗传改良提供理论依据。
1 材料与方法
1.1 试验材料
以贵州省册亨县油茶主产区弼佑乡(105.94°E,24.82°N)、秧坝镇(105.82°E,24.89°N)、冗度镇(105.73°E,25.04°N)、双江镇(106.09°E,24.86°N)的望谟红球油茶单株为试材,共计4个居群,每个居群选取8~14株无病虫害、生长正常、结实量稳定的成年植株,共43株,其中弼佑(P 1)11株,编号为1~11;秧坝(P 2)10株,编号为12~21;冗度(P 3)8株,编号为22~29;双江(P 4)14株,编号为30~43。每个单株从东、西、南、北4个方向均匀采集树冠中上部的果实。
1.2 果实性状测量
每个单株随机选取3组(30个/组)果实,用游标卡尺测定其果实纵径、横径、果皮厚度,并由纵径/横径的值得到果形指数;用天平测定茶籽鲜重,然后在105 ℃恒温下烘至恒重,称量并记录其干重,计算含水量。
表1 望谟红球油茶4个居群间表型性状的平均值、标准差及多重比较
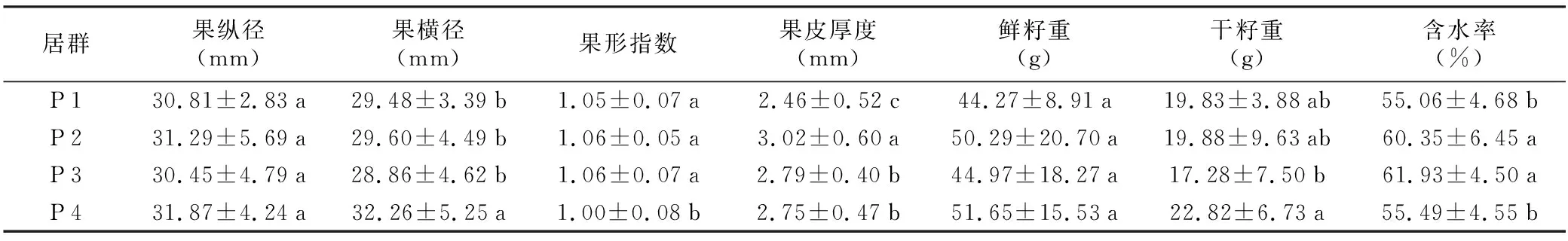
居群果纵径(mm)果横径(mm)果形指数果皮厚度(mm)鲜籽重(g)干籽重(g)含水率(%)P130.81±2.83a29.48±3.39b1.05±0.07a2.46±0.52c44.27±8.91a19.83±3.88ab55.06±4.68bP231.29±5.69a29.60±4.49b1.06±0.05a3.02±0.60a50.29±20.70a19.88±9.63ab60.35±6.45aP330.45±4.79a28.86±4.62b1.06±0.07a2.79±0.40b44.97±18.27a17.28±7.50b61.93±4.50aP431.87±4.24a32.26±5.25a1.00±0.08b2.75±0.47b51.65±15.53a22.82±6.73a55.49±4.55b
注:不同字母表示同一性状在不同群体间差异显著(p≤0.05),反之则差异不显著。
含水量(%)=(鲜籽重-干籽重)/鲜籽重×100%[4]。
1.3 数据统计分析
计算一般的描述性统计,包括均值、标准差和变异系数(CV)。计算各性状及分级的分布频率,采用Shannon-Wiener 指数表示形态多样性程度。计算不同性状的相关性,并根据平Euclidean距离按Ward 法(离差平方和法)进行聚类分析。最后通过主成分分析筛选决定遗传多样性的主要性状。利用Excel软件和SPSS 22.0软件进行数据统计和分析。
2 结果与分析
2.1 不同居群望谟红球油茶种实变异特征
对4个居群望谟红球油茶的果纵径(FVD)、果横径(FHD)等7个表型性状的平均值及标准差进行分析(表1)可知,4个居群的果纵径、鲜籽重2个性状无显著差异,P 4居群的果横径、果形指数2个性状与其他3个居群存在显著差异,各个性状的变异在居群间无显著规律。果皮厚度直接影响着望谟红球油茶的出籽率,果皮越薄,出籽率越高,从表1可以看出,P 1居群的果皮厚度最小,并与其他3个群体存在显著差异,而P 2居群的果皮厚度最大,也与其他3个居群存在显著差异。果形指数是划分果形的重要依据,P 4居群的果形指数与其它3个居群间存在显著差异,而其余3个居群间无显著差异,同时从表1可以看出,P 4居群的果实个体较大,果纵径、果横径和鲜籽重、干籽重均高于其他居群。茶籽含水量对油茶品质具有重要影响,福建小果油茶茶籽成熟后含水量平均值为55%[5],望谟红球油茶茶籽成熟后含水量略高,4个居群平均含水量均大于55%,其中P 1居群和P 4居群的茶籽含水率相对较低,且2个居群间无显著差异,但与另外2个居群存在显著差异,而P 2和P 3居群的含水率较高,达60%以上。
变异系数可以在一定程度上衡量性状的离散程度,变异系数越大,说明该种群的性状离散程度越大,表型多样性越高,反之,变异系数越小,性状离散程度越低[6]。从表2可以看出,望谟红球油茶种实各性状的平均变异系数(CV)为18.66%,变异幅度为6.78%~35.22%,其中果形指数(6.78%)和茶籽含水率(8.67%)的变异系数较小,干籽重(35.22%)和鲜籽重(33.00%)较大的变异系数较大,说明望谟红球油茶果形指数和茶籽含水率的变异幅度较小,稳定性高,而干籽重和鲜籽重的变异较大,稳定性差。3个籽粒相关性状的平均变异系数为25.63%,3个果形相关性状的平均变异系数为11.88%,说明望谟红球油茶果实的果形相关性状变异较小,籽粒相关性状变异较大。4个居群7个性状的平均变异系数为13.84%~22.68%,P 1居群的变异系数(13.84%)最低,该居群的表型多样性程度较低,P 2居群的变异系数(22.68%)最高,其表型多样性较为丰富。
表2 望谟红球油茶4个居群表型性状变异系数

表型性状居群P1P2P3P4平均果纵径(mm)9.2018.2015.7313.2914.11果横径(mm)11.5015.1816.0216.2814.75果形指数6.905.106.988.146.78果皮厚度(mm)21.0919.9914.3517.0418.12鲜籽重(g)20.1241.1740.6330.0733.00干籽重(g)19.5748.4543.3929.4735.22含水率(%)8.5010.697.278.208.67平均13.8422.6820.6217.5018.66
表3 望谟红球油茶果形和果色性状不同类型的频率分布和多样性指数
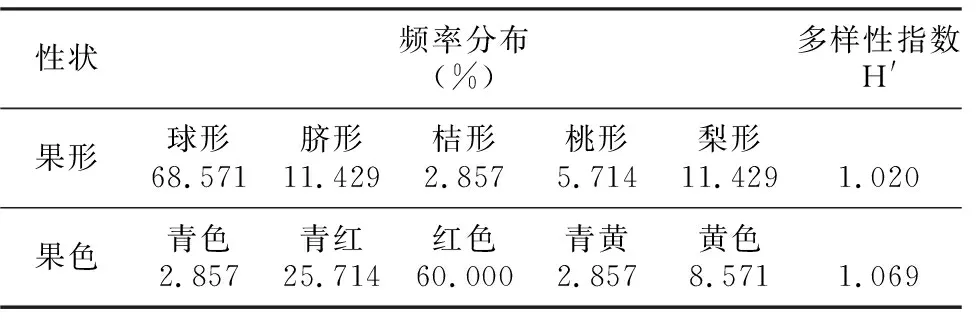
性状频率分布(%)多样性指数H'果形球形68.571脐形11.429桔形2.857桃形5.714梨形11.4291.020果色青色2.857青红25.714红色60.000青黄2.857黄色8.5711.069
2.2 望谟红球油茶种实性状的多样性分析
参照彭邵锋等[7]、姚小华等[8]的方法,根据果形指数和实际目测的效果,将参试的望谟红球油茶果形分为球形、脐形、桔形、桃形和梨形5类。望谟红球油茶的果形呈多样化,同一单株兼具有不同的果形。从表3可以看出,望谟红球油茶的果形主要为球形,占总数的68.571%,少数为桔形,仅占2.857%,其中,脐形是桔形的一个特殊变型;望谟红球油茶的颜色以红色为主,占总数的60%,少数为青色和青黄色。
表4 望谟红球油茶种实性状多样性指数
表5 望谟红球油茶种实性状的相关性分析
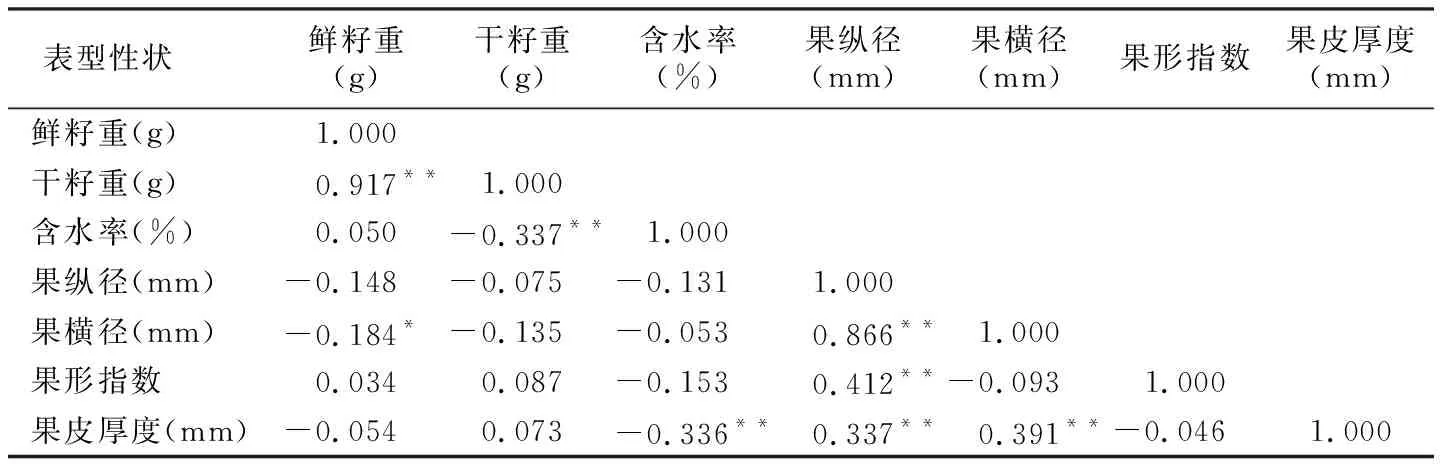
表型性状鲜籽重(g)干籽重(g)含水率(%)果纵径(mm)果横径(mm)果形指数果皮厚度(mm)鲜籽重(g)1.000干籽重(g)0.917**1.000含水率(%)0.050-0.337**1.000果纵径(mm)-0.148-0.075-0.1311.000果横径(mm)-0.184*-0.135-0.0530.866**1.000果形指数0.0340.087-0.1530.412**-0.0931.000果皮厚度(mm)-0.0540.073-0.336**0.337**0.391**-0.0461.000
注:“**”表示在0.01水平上极显著相关,“*”表示在0.05水平上显著相关。
Shannon-Weiner多样性指数是丰富度和均匀度的综合指标[9],通过计算4个居群望谟红球油茶7个种实性状Shannon-Weiner多样性指数(表4),结果表明,望谟红球油茶不同种实性状的多样性指数为1.923~2.080,变化范围小,均匀度高;7个种实性状的平均多样性指数值为2.021,变异丰富。
2.3 种实性状间的相关性分析
由表5可以看出,7个性状间存在不同程度的相关性,相关系数介于-0.337~0.917之间,但只有部分性状间存在显著或极显著相关关系。其中,果纵径与果横径和果形指数呈极显著正相关;干籽重和鲜籽重呈极显著正相关;果皮厚度与果纵径和果横径呈极显著正相关;干籽重和含水率呈极显著负相关;果横径和鲜籽重呈显著负相关;果皮厚度极显著的影响着含水率,二者呈极显著负相关关系。3个果形性状与3个籽粒性状间除了果横径与鲜籽重存在显著负相关关系外,其余各性状间无明显相关关系。
2.4 种实性状的主成分评价
采用主成分分析法对4个居群的43个望谟红球油茶单株7个种实性状进行综合分析(表6)表明,主成分构成的主要信息集中在前3个主成分,特征值总和为6.424,累计贡献率达91.775%,可以反映所有性状的大部分信息。主成分特征向量绝对值反映了其对主成分的影响程度[10],由表6可知,第一主成分特征值为3.694,贡献率52.773%,其中作用最大的性状有鲜籽重、干籽重、果纵径、果横径,其特征向量值分别为0.950、0.876、0.930、0.914;第二主成分特征值为1.591,贡献率22.731%,主要反映了含水率和果皮厚度2个性状,特征向量值分别为0.946和0.723;第三主成分特征值为1.139,贡献率16.271%,主要反映果形指数(0.980)。
表6 望谟红球油茶种实表型性状主成分分析

性状主成123鲜籽重(g)0.950-0.0220.132干籽重(g)0.876-0.3920.136含水率(%)0.0330.946-0.048果纵径(mm)0.9300.0080.210果横径(mm)0.914-0.064-0.303果形指数-0.1310.1260.980果皮厚度(mm)0.5540.723-0.060特征值3.6941.5911.139贡献率(%) 52.77322.73116.271累计贡献率(%)52.77375.50491.775
2.5 表型性状的聚类分析
基于7个种实表型性状指标对43个望谟红球油茶单株进行UPGMA聚类分析(图1)。结果显示,在欧氏距离15处可将43个望谟红球油茶单株划分为3个大类,第Ⅰ类包括9、23、14、……、4等20个单株;第Ⅱ类包括42、43、……、26等15个单株;第Ⅲ类仅有13、19、……、38等8个单株。而在欧氏距离6的阀值处每个大类又分别可以分为更小的2 个亚类。总的来看,43个单株并没有依据居群划分进行聚类,不同居群的望谟红球油茶单株分散在各大类中,并不存在区域上的特异性,可能由于各乡镇间存在引种的习惯,同时油茶为异花受粉植物,为油茶的变异提供了丰富的遗传基础。
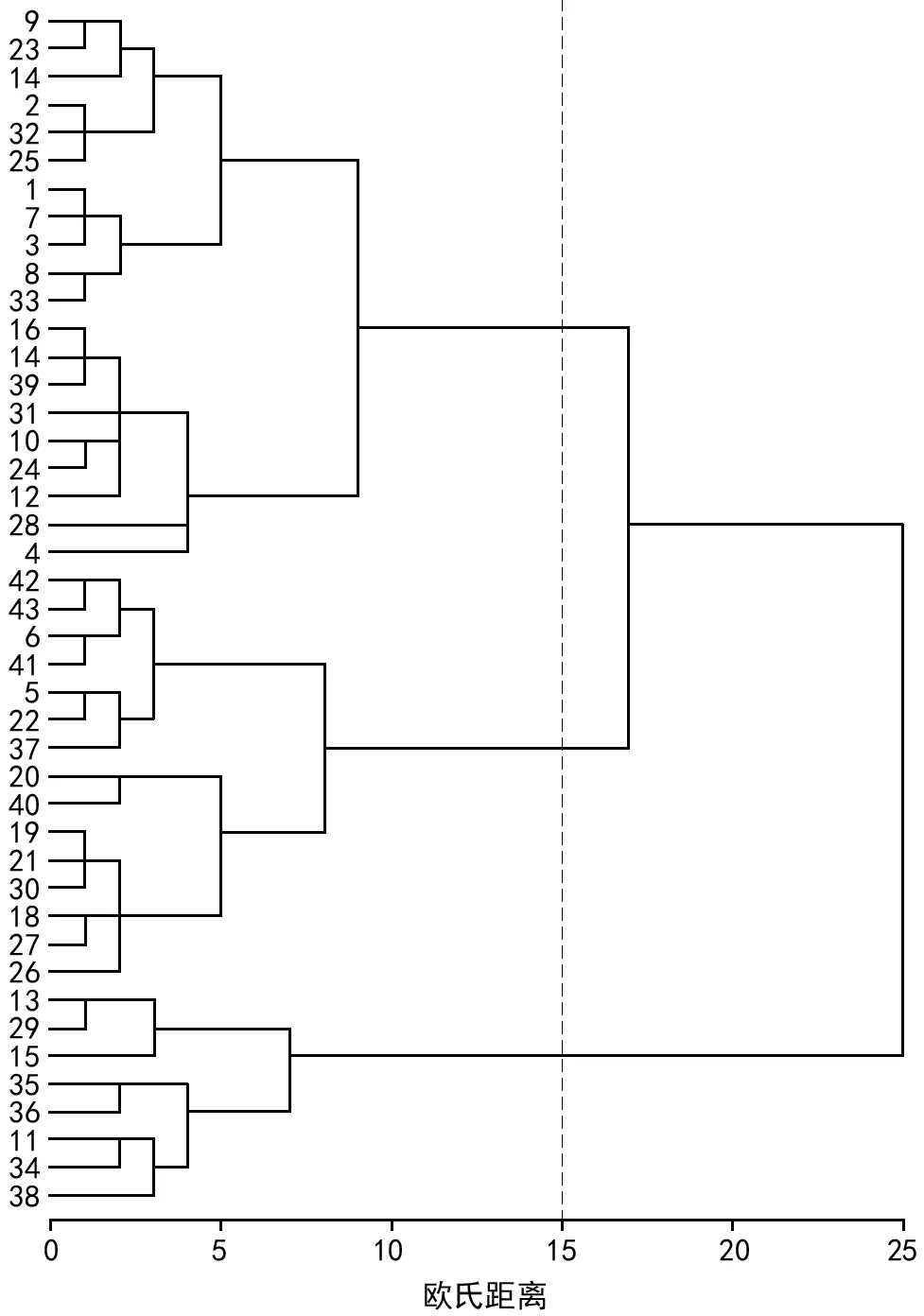
图1 43个望谟红球油茶单株表型性状聚类图
3 结论与讨论
表型多样性是遗传多样性与环境多样性的综合体现,既具有稳定性又具有变异性,是遗传多样性研究的重要内容[11-13],利用表型性状研究植物的遗传变异是最直接、简便易行的方法,因此通常依据植物表型性状进行种植资源的分类、鉴定及育种材料选择[14-17]。通过对册亨县望谟红球油茶4个居群43个单株的7个表型性状的变异进行分析发现,望谟红球油茶种实性状变异丰富,4个居群7个性状的平均变异系数均高于10%,其中P 1居群的变异系数(13.84%)最低,而P 2居群的变异系数(22.68%)最高。从差异显著性结果看:4个居群不同性状间具有不同程度的差异性。4个居群望谟红球油茶7个种实性状Shannon-Weiner多样性指数为1.923~2.080,平均多样性指数为2.021,变异丰富。目前,望谟红球油茶多处于野生、半野生和实生栽培状态,资源丰富、天然杂交形成的自然变异和人工选择的干预,形成了多种变异类型。广泛的变异和丰富的遗传基础,可为望谟红球油茶遗传改良提供丰富的材料。
对望谟红球油茶果形和果色性状的频率分布和多样性指数分析发现,望谟红球油茶的果形呈多样化,主要为球形、脐形、桔形、桃形和梨形5大类,但球形分布频率最大(占总数的68.571%);果色以红色为主(占总数的60%),少数为青色和青黄色。谢一青等对小果油茶果实性状与含油率及脂肪酸组成的相关性进行分析,结果表明:小果油茶籽含油率与果形有关,种仁含油率和干籽含油率顺序均为:脐形果>球形果>桃形果;油酸含量的顺序为桃形果>脐形果>球形果,而多不饱和脂肪酸含量的顺序为桃形果<脐形果<球形果[18],康志雄等[19]对普通油茶进行研究也得到相似的结论。由于果形是特征明显、易于辨别、遗传稳定性较高的质量性状,因此在油茶新品种选育中,可首先通过果形对油茶籽油的品质进行间接选择,在此基础上,再选择高产品种,以减少选择育种的工作量。
主成分分析是将多个变量转化为少数几个综合因子,每个主成分基本上又能反映原始变量的大部分信息,其所含信息互不重复,依据各因子的贡献率大小确定其重要性[20]。从主成分分析结果可知,前3个主成分的累计贡献率达91.775%,基本反映了所有43个单株7个表型性状的大部分信息。第一主成分特征向量较大的性状有鲜籽重、干籽重、果纵径、果横径,反映了果实大小和茶籽重量具有较高的育种选择潜力。第二主成分主要反映了含水率和果皮厚度2个性状,茶籽含水量与种子品质有关,因此,在望谟红球油茶优良单株或品种的筛选中,含水量可作为一个重要的参考指标。第三主成分反映了果形指数在育种选择中也占有一定地位。
望谟红球油茶4个居群43个单株表型性状的聚类结果显示,43个单株并没有完全依据居群划分进行聚类,没有区域上的特异性,可能是因为望谟红球油茶为异花授粉植物,栽培历史悠久,具有较为广泛的种内杂交,而且各乡镇间存在相互引种的习惯。由于表型性状容易受到自然环境和人为因素的影响[21],仅凭表型性状难以准确把握望谟红球油茶居群及单株间的亲缘关系,要深入了解望谟红球油茶的遗传多样性,仍需利用SSR、ISSR、SRAP、RAPD、AFLP 等分子标记技术对其进行更深入的研究,同时结合表型多样性的研究,为望谟红球油茶资源的保护和利用提供理论依据。
参考文献:
[1]陈晓春.望谟红球油茶性状评价与发展[J].贵州林业科技,1992,20(3):43-47.
[2]罗在柒,王进,邱建生,等.望谟红球油茶林产量与种群构件特性调查研究[J].贵州林业科技,2011,39(4):23-27.
[3]黄勇,姚小华,王开良,等.小果油茶种实表型性状遗传多样性研究[J].安徽农业大学学报,2011,38(5):698-707.
[4]沈柳杨,张宏,范修文,等.含水率对温185核桃静压力学性能的影响[J].山东农业大学学报(自然科学版),2016(3):332-337.
[5]郑德勇,凌宏有.福建油茶籽在成熟过程中重要成分动态变化的研究[J].中国粮油学报,2014,29(2):36-43.
[6]冯秋红,史作民,徐峥静茹,等.岷江柏天然种群种实表型变异特征[J].应用生态学报,2017,28(3):748-756.
[7]彭邵锋,陈永忠,张日清,等.油茶果形果色分类及经济性状[J].中南林业科技大学学报,2007,27(5):33-39.
[8]姚小华,黄勇.小果油茶资源与遗传多样性研究[M].北京:科学出版社,2013:14.
[9]黄勇,姚小华,王开良,等.小果油茶种实形态变异频率及其多样性指数分析[J].江西农业大学学报,2011,33(2):292-299.
[10]王楚天,游璐,钟琳珊,等.江西29个油茶品种始果早期表型变异分析[J].西南林业大学学报,2017,37(1):73-81.
[11]Pigliucci M,Murren C J,Schlichting C D.Phenotypic plasticity and evolution by genetic assimilation[J].Journal of Experimental Biology,2006,209(12):2 362-2 367.
[12]Lande R.Adaptation to an extraordinary environment by evolution of phenotypic plasticity and genetic assimilation[J].Journal of Evolutionary Biology,2009,22(8):1 435-1 446.
[13]谷衍川,丰震,李承水,等.小紫珠种群的表型遗传多样性研究[J].山东林业科技,2013,43(2):5-9.
[14]王永康,吴国良,赵爱玲,等.枣种质资源的表型遗传多样性[J].林业科学,2014(10):33-41.
[15]丹曲,张艳福,方江平,等.西藏林芝不同居群桃儿七表型多样性分析[J].南方农业学报,2017,48(3):386-392.
[16]邓丽丽,朱霞,和润喜,等.云南松不同茎干类型种实性状表型多样性比较[J].种子,2017,36(3):4-9.
[17]张晓骁,宋超,张延龙,等.秦岭与子午岭地区紫斑牡丹居群表型多样性研究[J].园艺学报,2017,44(1):139-150.
[18]谢一青,李志真,姚小华,等.小果油茶果实性状与含油率及脂肪酸组成相关性分析[J].中国油脂,2013,38(5):80-83.
[19]康志雄,邹达明.油茶经济性状通径分析和聚类分析研究[J].福建林学院学报,1989,9(2):145-151.
[20]魏忠芬,李慧琳,奉斌,等.贵州紫苏种质资源表型性状的遗传多样性[J].西南农业学报,2017,30(1):45-52.
[21]尹晓蛟,万志能,袁伟,等.麻城山茶种质资源表型遗传多样性研究[J].江西农业学报,2017,29(3):62-65.
