受母性影响的油菜种子性状研究
2018-06-08,,,,,,,
, , , , , , ,
(1.云南农业大学农学与生物技术学院, 昆明 650201;2.云南省农业科学院园艺作物研究所, 昆明 650205;3.云南农业大学社会服务办公室, 昆明 650201; 4.云南农业大学农科教学实验中心, 昆明 650201)
油菜是四大主要油料作物之一,在世界油料作物中发挥着举足轻重的作用,油菜籽全球总产仅次于大豆[1],同时油菜也是植物蛋白质饲料来源[2]。研究表明,作物正反交种子的某些性状会产生差异,是由于受母体植株基因型的影响和卵细胞的细胞质效应。E.Millet等在研究小麦正反杂交过程中发现,其籽粒蛋白质的含量受母体效应的影响[3]。Singh等认为,大豆籽粒蛋白质含量具有较强的母体效应[4]。Brim等研究发现,大豆籽粒油分含量也具有较强的母体效应[5]。D.G.Dan等在棉花杂交中发现,种子的含油量存在母体效应,而且母体效应高于基因遗传效应[6]。目前,关于油菜种子性状的遗传控制研究还较少,仅王新发[7]对甘蓝型油菜种子含油量进行了研究。为此,本研究选取种子含油量、大小等性状差异显著的2个甘蓝型油菜DH系进行正反交,并对正反交种子进行性状比较分析,旨在探索油菜种子性状的遗传控制机制,为解析其分子机制提供理论指导和依据。
1 材料与方法
1.1 材 料
亲本材料为甘蓝型油菜(BrassicanapusL.)品系DH 0729和DH 0216,均由云南农业大学油菜研究室提供。
1.2 方 法
1.2.1 材料种植
为了使2个亲本花期相遇,进行不同时期播种,种植在云南农业大学后山农场试验地。2016年10月5日播种DH 0729,2016年10月15日播种DH 0216,后期进行油菜常规栽培管理。
1.2.2 人工去雄与授粉
选择生长健壮无病虫害的植株为试验对象。父本于授粉前3 d摘除已开放的花蕾,并套袋隔离。于授粉前1 d摘除母本主花序上已开放花蕾及上部较小的花蕾,保留5~7个较大的将要开放的花蕾进行人工去雄,然后套袋隔离,并挂牌标记。于次日09:00—11:00时用棉签将父本花粉授在母本柱头上,并立即套袋隔离。以DH 0729为母本,DH 0216为父本配制正交组合,记为DH 29-16。以DH 0216为母本,DH 0729为父本配制反交组合,记为DH 16-29。成熟时收获种子,并统计角果数。
1.2.3 种子形态性状的测定
种子大小以千粒重、直径计量。用千分之一电子分析天平随机称量1 000粒种子的重量,重复3次。用电子游标卡尺测量种子直径,每次测量50粒种子,重复3次。在体视显微镜下将种皮、子叶、胚剥离开,每次50粒种子,然后分别称量种皮、子叶、胚的重量,重复3次。
1.2.4 含油量的测定方法
将除去杂质干净的油菜种子寄到河南大学河南省植物逆境生物学重点实验室,用丹麦 Foss 公司NIR 5000型近红外光谱分析仪测定种子的含油量和几种常见脂肪酸的含量及硫甙的含量。
1.2.5 种子不同组织的含油量测定
分别称取0.2 g种皮、子叶、胚放入玻璃离心管中,在65 ℃烘箱中处理1周,完全烘干脱水后转入研钵中,加液氮充分研磨成粉末,然后将粉末全部转移至玻璃提油管中,加800μL的正己烷/异丙醇(3∶2)提取液,静置1 min后在涡旋仪上振荡混匀10 min,然后3 000 r/min离心8 min,将上清转移到另一已称重的提油管中,并再在沉淀中加入500μL正己烷/异丙醇(3∶2)提取液,在涡旋仪上振荡混匀10 min,3 000 r/min离心10 min,转移上清并合并两上清液,真空干燥,然后称重,减去管重即获得油脂重量。每种组织重复测定3次。
2 结果与分析
2.1 油菜正反交结角果率的差异
对油菜正反交的结角果率进行分析,结果(表1)表明,正反交的结角果率有一定的差异,正交的结角果率为85.9%,反交的结角果率为77.1%,反交的结角果率要低于正交的结角率。理论上同一物种不同品种或品系间杂交不会存在授粉和结实的差异,但在本研究中正反交结角果率表现出差异,可能是由于在人工授粉去雄过程有一些花蕾的柱头或子房受到了损伤,使授粉到柱头上花粉不能很好地萌发和受精,导致角果发育不正常而脱落。
表1 正反杂交组合结角果率

杂交组合授粉花蕾总数(个)成熟角果数(个)结角果率(%)DH29-1652144885.9DH16-2958645277.1
2.2 油菜正反交种子大小的差异
对亲本以及正反交种子千粒重进行分析,结果(图1)表明,亲本千粒重差异显著,亲本DH 0729的千粒重为5.074 0 g,亲本DH 0216的千粒重为3.301 1 g。但其正反交种子千粒重差异不明显,正交种子千粒重为6.060 4 g ,反交种子千粒重为5.998 9 g。对亲本及正反交种子的直径进行分析,结果(图2)也表现出同样的规律。2个亲本的种子直径差异明显,DH 0729
表2 种皮、子叶、胚占种子的占比(单位:%)
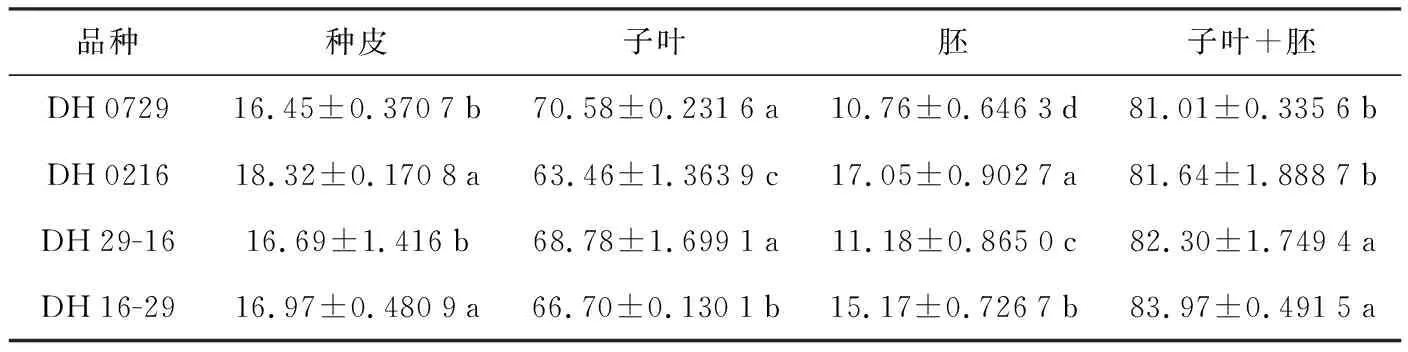
品种种皮子叶胚子叶+胚DH072916.45±0.3707b70.58±0.2316a10.76±0.6463d81.01±0.3356bDH021618.32±0.1708a63.46±1.3639c17.05±0.9027a81.64±1.8887bDH29-1616.69±1.416b68.78±1.6991a11.18±0.8650c82.30±1.7494aDH16-2916.97±0.4809a66.70±0.1301b15.17±0.7267b83.97±0.4915a
表3 种子各种脂肪酸差异分析(单位:%)
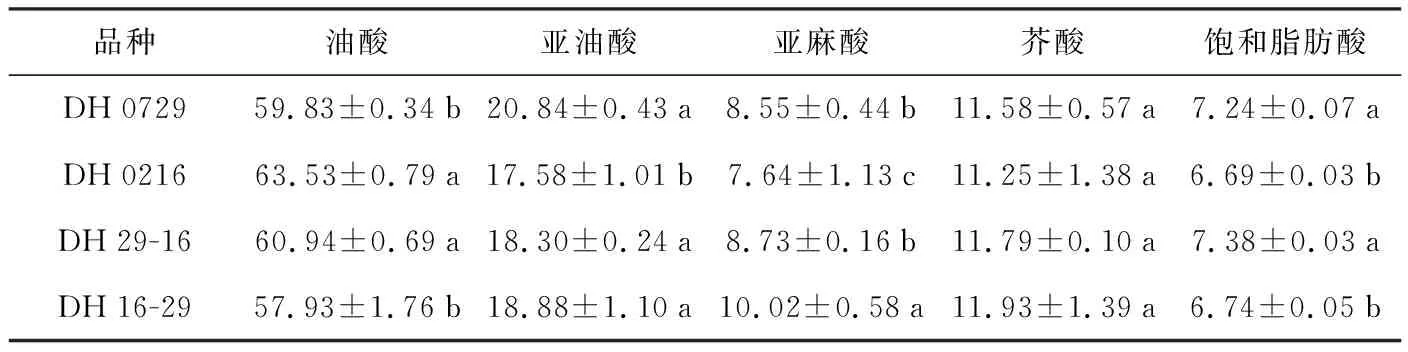
品种油酸亚油酸亚麻酸芥酸饱和脂肪酸DH072959.83±0.34b20.84±0.43a8.55±0.44b11.58±0.57a7.24±0.07aDH021663.53±0.79a17.58±1.01b7.64±1.13c11.25±1.38a6.69±0.03bDH29-1660.94±0.69a18.30±0.24a8.73±0.16b11.79±0.10a7.38±0.03aDH16-2957.93±1.76b18.88±1.10a10.02±0.58a11.93±1.39a6.74±0.05b
表4 种子不同组织含油量分析(单位:%)

品种种子含油量种皮子叶胚DH072937.47±0.76b2.87±0.10b33.65±2.68b36.30 ±1.79aDH021646.20±1.16a1.91±0.15c42.40±2.35a37.06±1.32aDH29-1638.11±0.22b2.70±0.17a35.42±1.71b34.75 ±1.72aDH16-2946.66±1.86a2.15±0.15c42.15 ±1.75a36.77±1.44a
种子的平均直径为2.005 9 mm,属于大种子,DH 0216种子的平均直径为1.221 5 mm,属于小种子。而正反交种子直径的差异不显著,正交种子的平均直径为2.260 1 mm,反交种子的平均直径为2.136 9 mm。由此可见,不论是正交,还是反交,种子大小并未表现出偏向性,不受母体效应(基因型)的影响。正反交种子的大小均大于最大的亲本,并且差异显著,这是由于在进行杂交时每个花序只保留了5~7朵花,有充足的营养供应角果和种子发育之需要。另外,正反交杂种子的成熟期延缓,增加了种子发育和灌浆的时间,从而使正反交种子的千粒重和直径显著地增加。
2.3 油菜种子不同组织的占比分析
对油菜种子不同组织的占比进行分析,结果(表2)表明,在油菜种子中,子叶占比在60%以上,而种皮和胚的占种比较小,均在20%以下。种皮、子叶以及叶+胚的占比在亲本和正反交种子中差异都不明显,而且正反交的也没明显地表现出偏母性。但胚的占比在2亲本之间及正反交之间差异显著,而且表现出明显的偏母性,母本占比低的其后代的占比也低,母本占比高的其后代得占比也较高,说明胚的占比存在母体效应。
2.4 种子含油量与脂肪酸组成的差异分析
利用近红外仪对亲本和正反交种子的含油量进行测定,发现正反交种子的含油量差异显著,母本含油量高的,其杂交种子的含油量也高;母本含油量低的,其杂交种子含油量也低,说明油菜种子含油量存在明显母体效应,具有偏母性。采用重力法对种子不同组织的含油量进行测定,发现种皮的含油量普遍较低,为2%左右,而子叶和胚的含油量较高,均在30%以上,说明油脂主要在子叶和胚中累积。此外,正反交种子的种皮、胚的含油量差异不明显,没有呈显出偏母性,而正反交种子的子叶含油量差异显著,并呈现出偏母性。这说明正反交种子含油量的偏母性是由子叶含油量的偏母性决定的,是由于子叶的含油量比较高,特别是子叶在种子中的占比表现特别高,达到70%。虽然胚的含油量较高,但由于其在种子中的占比表现特别低,不到20%,因此,不会影响到种子含油量的偏母性。但在本研究中所测得的油菜种子不同组织的含油量都低于整个种子含油量,这主要与测定方法有关,因为在组织含油量的测定过程中有一定的油脂损失,如组织粉末转移不完全或油脂抽提不完全等。

图1 种子千粒重显著性比较
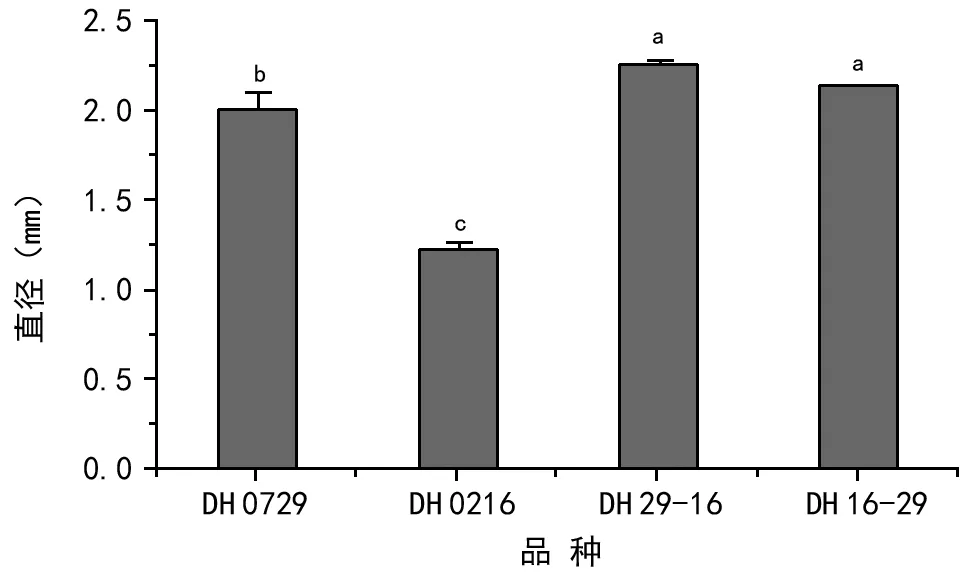
图2 种子的直径显著性分析
利用近红外仪测定了亲本和正反交种子几种主要脂肪酸的含量,发现油酸、亚麻酸、饱和脂肪酸的含量在亲本间存在一定的差异,在正反交种子间同样也存在一定的差异,但并未呈现出偏母性。而亚油酸含量在亲本间存在一定的差异、但在正反交种子间没有差异。仅芥酸含量在亲本、正反交种子间差异不明显,这可能与它们均为双低油菜、其芥酸基因隐性纯合有关。
3 讨 论
油菜种子性状的遗传机制研究,有利于遗传高产育种。本研究发现,正反交种子千粒重、直径都显著大于亲本。王杰利在对甘蓝型营养分配研究过程中发现,适当的营养分配调控可以使油菜主花序角果收获的种子千粒重增加,最高可以增加1倍左右[8],本研究结果与之相符。杨光等研究发现,油菜正反杂交后能导致种子胚的干重略微增加[9]。本研究也发现,正反杂交子叶和胚总重比均略大于亲本,但是胚/种比具有明显偏母性,而种皮和子叶占比无明显的偏母性,而前人未见研究和报道。同时对油菜种子的组分分析发现,油菜种子大小差别主要来自于子叶占整个种子的比,子叶占种比越大种子就越大。
本研究发现,正反交种子及组分中子叶含油量具有明显偏母性,而种皮、胚的含油量未表现出偏母性。韦存虚等在对油菜种子不同组织的油体进行观察发现,种子子叶细胞大部分区域都被油体所充实[10]。本研究也发现,油菜的油脂主要储存在子叶中。对种子不同组织的占比和含油量分析发现,种子中子叶/种比远远大于胚/种比,虽然子叶和胚在种子中含油量相差不大,但是胚/种比明显低于子叶/种比,说明油菜正反交种子含油量高低主要是其子叶大小和含油率共同决定。但是目前对于油菜正反交种子含油量偏母性的调控机制还不清楚。王通强在对甘蓝型油菜的杂交后代种子含油量的遗传做了部分研究,发现后代种子含油量与母体含油量呈显著正相关 , 而与父本含油量则无明显相关性[11]。同时油菜种子含油量属于多基因控制的数量性状,较易受到外界环境因素的影响,导致含油量差异性。作为油料作物,油菜种子在发育过程中,物质的积累比较复杂,涉及糖和脂肪累积2个不同阶段,在中间代谢方面会受到大量调节因素的控制,使得含油量遗传调控变得极其复杂[12],因此对含油量调控解析进展缓慢,对于正反交导致含油量的差异和偏母性原因至今没有很好的解释。
种子含油量大小由种子千粒重和含油率构成。因此,在以后油菜育种过程中,可以通过植株角果合理的营养分配提高种子千粒重,在分子水平对子叶含油率进行调控,最终达到增加油菜种子产油量的目的,从而培育出超高产油菜新品种。
参考文献:
[1]郑循刚,罗怀凤,傅新红.四川省油菜产业发展研究[J].农业技术经济,2002(1):54-56.
[2]李娜,杨涛.国家扶持政策对我国油菜籽产业发展的影响[J].粮油加工,2009(1):22-25.
[3]E.Millet,周恒.小麦粒重和籽粒蛋白质含量的父、母体效应[J].麦类作物学报,1992(5):7-10.
[4]Singh,L.,Hadley,H.H.Maternal and Cytoplasmic Effects on Seed Protein Content in Soybeans,Glycinemax(L.)[J].Merrill.Cropence,1972,12(5):583-585.
[5]Brim,C.A.,Schutz,W.M.,Collins,F.I.Maternal Effect on Fatty Acid Composition and Oil Content of Soybeans,Glycinemax(L.)[J].Merrill.Crop Science,1968(5):517-518.
[6]Dani,R.G.,Kohel,R.J.Maternal effects and generation mean analysis of seed-oil content in cotton (GossypiumhirsutumL.)[J].Theoretical & Applied Genetics1989,77(4):569.
[7]王新发.甘蓝型油菜含油量的遗传与生态学研究及其杂种优势利用[M].北京:中国农业科学院,2011.
[8]王杰利.营养分配调节对甘蓝型油菜种子大小的影响及其机制[J].江苏农业学报,2016,32(4):759-762.
[9]杨光,孙统庆,左青松,等.油菜杂交种和亲本胚的增重特性差异[J].广东农业科学,2013,40(5):15-17.
[10]韦存虚,钦风凌,李爱民.油菜种子油体的观察和大小分析[J].中国油料作物学报,2009,31(4):445-448.
[11]王通强.油菜籽含油量的遗传及杂种优势[J].贵州农业科学,1992(6):37-40.
[12]Baud S,Lepiniec L.Physiological and developmental regulation of seed oil production[J].Progress in Lipid Research,2010,49(3):235-249.
