从T7噬菌体展示cDNA文库筛选生殖支原体黏附蛋白的互作蛋白①
2018-05-25邓湘赢余敏君李玲玲曾焱华
戴 佩 邓湘赢 余敏君 李玲玲 罗 丹 龙 娴 曾焱华
(南华大学病原生物学研究所,特殊病原体防控湖南省重点实验室,湖南省分子靶标新药研究协同创新中心,衡阳 421001)
生殖支原体(Mycoplasma genitalium,Mg)是一种能在无生命培养基中生长的最小原核细胞型微生物,与很多泌尿生殖道感染性疾病有关,如男性和女性不良生殖后遗症、子宫内膜炎、盆腔炎症性疾病、宫颈炎和非淋菌性尿道炎[1]。研究表明生殖支原体可能是人类免疫缺陷病毒(Human immunodefici-ency virus,HIV)的协同因子,能促进HIV的感染和复制[2,3]。此外,生殖支原体可与人类精子结合,并随活动的精子引起扩散性感染,是引起女性生殖系统疾病和不孕不育的一个重要因素[4]。
生殖支原体黏附蛋白(MgPa)是暴露在Mg膜表面的主要黏附蛋白,具有较强的抗原性和免疫原性,与Mg感染或侵入宿主细胞并进一步引起宿主细胞受损和死亡有着紧密的联系[5]。Mg感染或侵入宿主细胞的过程,其本质是Mg与宿主细胞发生相互作用的过程,鉴于目前国内外尚未见筛选与鉴定MgPa互作蛋白的报道,本课题组在前期研究中表达并纯化了重组MgPa(rMgPa),制备了纯化的抗rMgPa的抗体[6],且构建了人尿道上皮细胞T7噬菌体展示cDNA文库[7],本研究拟以rMgPa为靶分子,从人尿道上皮细胞(SV-HUC-1)T7噬菌体展示cDNA文库中筛选MgPa的互作蛋白,为进一步了解MgPa的功能、生殖支原体的致病机制及其防治提供实验依据。
1 材料与方法
1.1实验材料、试剂 rMgPa的表达与纯化参照本课题组前期研究[5];抗rMgPa抗体由本课题组前期制备并保存;SV-HUC-1细胞T7 噬菌体展示cDNA文库由本课题组前期构建[7];BLT5403菌株由南华大学病原生物学研究所保存;T7筛选前引物和T7筛选后引物均购自美国No-vagen公司;HRP标记的T7-tag抗体购自中国博奥森公司;RPL35(ab121244)抗体购自Abcam公司;SV-HUC-1细胞购自中国科学院上海细胞库。
1.2方法
1.2.1SV-HUC-1细胞T7噬菌体展示cDNA文库的亲和筛选 以100 μg/ml的rMgPa蛋白包被96孔板,同时设立复孔和空白对照孔, 4℃湿盒过夜。经封阻液(BSA)于4℃封闭2 h。弃除封阻液,PBST洗6~8次。加入200 μl扩增后的噬菌体文库,滴度为3×108pfu[8],室温平摇2 h。倾倒未结合的噬菌体,甩干后用PBST洗6~8次,加150 μl的高盐洗脱液[9],室温平摇30 min,收集裂解液于4℃保存备用。将100 μl第1轮筛选后的噬菌体洗脱液加入预先包被rMgPa的微孔板,于37℃孵育2 h,弃孔内液,1×PBST(0.3% Tween-20)洗板6~8次,加入100 μl BLT5403菌液,37℃孵育2 h,按上述方法得到第2轮噬菌体洗脱液。按第2轮筛选方法再筛选2轮,取200 μl第2轮筛选后的洗脱液与空板孵育2 h,同时用含0.5% Tween-20的PBST洗板以排除与ELISA板结合的非特异噬菌体。第4轮筛选时将rMgPa的浓度降低为50 μg/ml以增强筛选的特异性。进行4轮筛选之后,即得到与rMgPa高亲和的噬菌体。
1.2.2噬菌斑的PCR扩增 挑选阳性噬菌斑进行PCR扩增,PCR引物为T7 筛选前引物:5′-GGAGCTGTCGTATTCCAGTC-3′和T7 筛选后引物:5′-AACCCCTCAAGACCCGTTTA-3′。PCR扩增条件如下: 94℃ 10 min,94℃ 50 s,50℃ 1 min,72℃ 1 min,30个循环,72℃延伸10 min。扩增产物经1.5%琼脂糖凝胶电泳分析后送上海生工生物有限公司进行DNA测序,并将其DNA序列进行BLAST分析。
1.2.3ELISA检测rMgPa与阳性噬菌体的特异结合 用100 μl rMgPa(100 μg/ml)包被ELISA板,同时设立复孔和阴性对照孔(WT型噬菌体),4℃湿盒过夜。弃掉包被液,加满封阻液(5%脱脂奶粉)于4℃封阻2 h,用PBST洗8次后拍干,加150 μl阳性噬菌体,37℃孵育2 h后用 PBST洗8次,加抗T7抗体(1∶1 000)于37℃孵育2 h,PBST洗8次后加入HRP标记的羊抗兔IgG抗体(1∶4 000),37℃孵育1 h 后用PBST洗8次,加A、B显色液,37℃避光孵育15 min后立即加入终止液,用酶标仪在450 nm处测吸光度值(A450)。
1.2.4斑点免疫印迹法验证rMgPa与阳性噬菌体的特异结合 用甲醇浸泡PVDF膜至透明后用TBST(0.5% Tween20)清洗,取1 μl噬菌体(107pfu)滴加在PVDF膜上,同时设立BSA对照、空白对照和阳性对照(rMgPa),待室温干燥后4℃封闭过夜。用TBST洗膜3次后,加入1 ml rMgPa(1∶100)于37℃孵育2 h,用TBST洗膜6次后再加入抗rMgPa的抗体(1∶1 000),37℃孵育2 h后洗膜6次,加入HRP标记的羊抗兔IgG抗体(1∶4 000)于37℃浸膜1 h,用TBST洗膜6次,ECL 法发光、显影。
1.2.5SV-HUC-1的培养与细胞总蛋白的提取 用含10% 胎牛血清和1%双抗(抗青霉素和链霉素)的F-12K培养基,在37℃、5% CO2的条件下培养SV-HUC-1,待细胞密度达90%时用PBS洗涤细胞3次,经胰酶消化后收集约5×107个细胞。1 000 r/min离心6 min,弃掉培养基。加1 ml经冰浴的PBS以重悬沉淀,4℃ 1 000 r/min离心5 min,重复3次。弃上清后加细胞裂解液500 μl以溶解细胞,冰浴25 min后超声处理5 min,加100 μl PBS重悬,即为细胞总蛋白,保存在-20℃备用。
1.2.6Far-western blot验证rMgPa与RPL35的特异结合 将rMgPa经12%SDS-PAGE胶分离。蛋白条带经半干转印系统转印到PV-DF膜上。用5%脱脂牛奶封闭过夜,TBST洗膜5次,加入提取的SV-HUC-1细胞总蛋白于4℃孵育6~10 h[10],TBST(0.5%Tween20)洗膜5次,加RPL35抗体(1∶200),4℃孵育过夜,TBST洗膜5次,加HRP标记的羊抗兔IgG抗体(1∶2 000)于37℃孵育1 h,TBST洗膜后ECL 法发光、显影。
2 结果
2.1与rMgPa特异结合的噬菌体得到明显富集 为筛选到与rMgPa相互作用的噬菌体,本实验以rMgPa为靶分子,通过“结合-洗脱-扩增”步骤对 SV-HUC-1的T7噬菌体展示cDNA文库进行了4轮亲和筛选,每次噬菌体的投入量保持一致(3×108pfu),如表1所示:第1轮淘选后的产率为0.67×10-4,而第4轮淘选时其产率达到了1.7×10-3,表明与rMgPa特异结合的噬菌体明显得到了富集。
2.2成功扩增了噬菌体的外源基因 为扩增噬菌体含有的外源基因,对从第4轮筛选的平板上随机挑取的32个噬菌斑进行PCR扩增,PCR产物采用1.5%琼脂糖凝胶电泳分析,结果显示成功提取到32个噬菌体的外源DNA,其片段大小介于250~750 bp之间,且500 bp左右的克隆最多(图1)。
2.3序列比对与同源性分析 为分析噬菌体含有的外源基因序列,将图2中PCR扩增的噬菌斑阳性产物送上海生工生物工程有限公司进行DNA测序。将测序后的序列在NCBI基因数据库中进行BLAST比对,结果表明32个噬菌体的外源序列中,共包括7种不同的序列,分别用P1~P7表示,其中RPL35蛋白出现的频率最高,达到了62.5%,结果如表2所示。
表14轮亲和筛选投入量和产出量的比率
Tab.1Ratioofoutputandinputoffourroundsbyaffinityscreening

Rounds1234Input(pfu)3×1083×1083×1083×108Output(pfu)2×10442×10419×1055×105Ratio067×10-414×10-463×10-417×10-3
2.4ELISA检测阳性噬菌体与rMgPa的特异结合 将上述7种代表性噬菌体包被在96板中, 采用间接ELISA法检测代表性噬菌体与rMgPa的特异结合,如图2所示:rMgPa组的A450值明显高于 BSA组,且P1、P2和P3与MgPa结合力较强。
2.5斑点免疫印迹检测阳性噬菌体与rMgPa的特异结合 采用斑点免疫印迹法进一步检测7个代表性噬菌体能否与rMgPa特异结合,结果如图3所示:P1、P2和P3代表性噬菌体出现了明显的斑点,说明P1(RPL-35)、P2(Mitochondrio complete genome)和P3(RPN4)能与rMgPa特异结合,这些结果与上述ELISA的结果一致。
2.6RPL35能与rMgPa特异结合 采用Far-western blot方法进一步检测SV-HUC-1细胞总蛋白中RPL35与 rMgPa的特异结合情况,rMgPa蛋白用SDS-PAGE分离并转至PVDF膜上,用提取的SV-HUC-1细胞总蛋白孵育后,用相应抗体检测,结果显示在约37 kD处有明显条带,说明RPL35能与rMgPa特异结合(图4)。
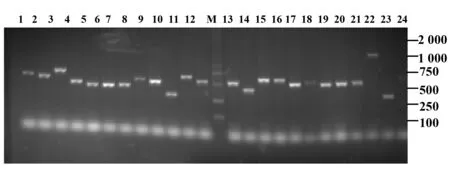
图1 部分噬菌斑PCR产物的琼脂糖凝胶电泳分析Fig.1 Agarose gel electrophoresis analysis of PCR products of partial phagesNote:M.DNA marker;Lane 1-24.Number of different phage plaques.
表2噬菌斑PCR产物同源性分析结果
Tab.2ResultsofhomologyanalysisofPCRproductsofphageplaque
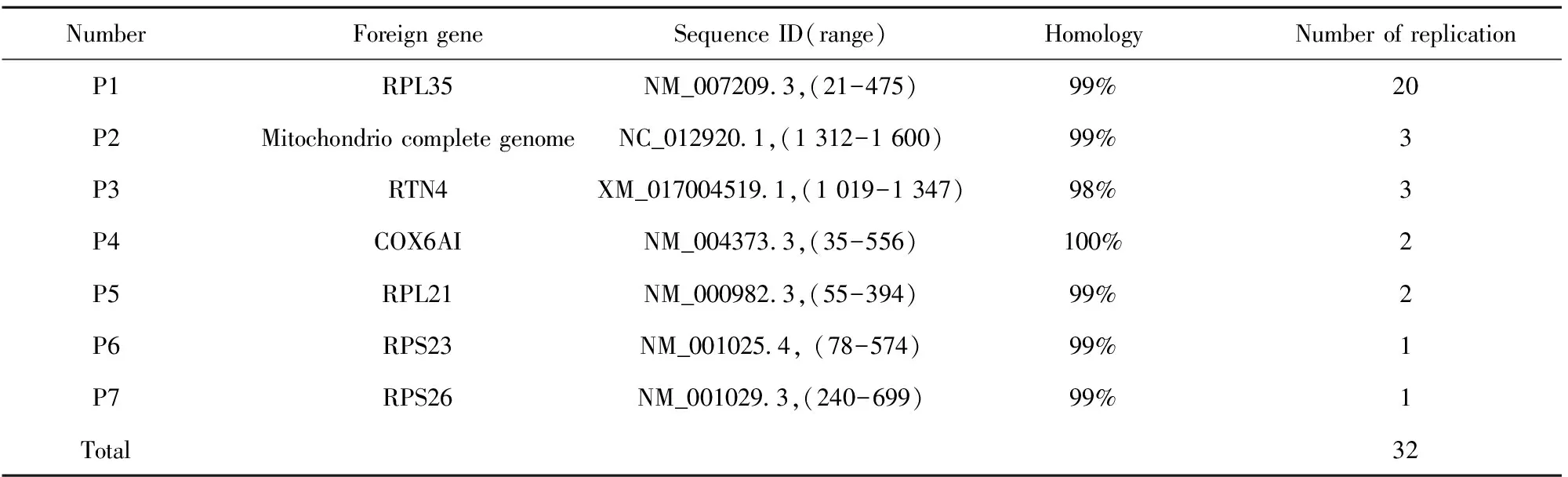
NumberForeigngeneSequenceID(range)HomologyNumberofreplicationP1RPL35NM_0072093,(21-475)99%20P2MitochondriocompletegenomeNC_0129201,(1312-1600)99%3P3RTN4XM_0170045191,(1019-1347)98%3P4COX6AINM_0043733,(35-556)100%2P5RPL21NM_0009823,(55-394)99%2P6RPS23NM_0010254,(78-574)99%1P7RPS26NM_0010293,(240-699)99%1Total32
Note:Homological analysis results to the inserted sequences of thirty-two phages,and the seven proteins edcoded by these genes were numbered as P1 to P7 as shown in table.
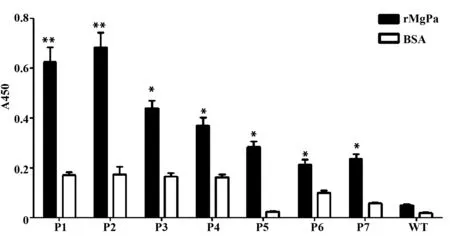
图2 ELISA检测代表性噬菌体与rMgPa的特异结合Fig.2 Specific combination between representative phages and rMgPa was detected by ELISANote:P1-P7.Number of representative phages;WT.Wild type phage.BSA was used as blank control.Compared with the BSA group,**.P<0.01;compared with the BSA group,*.P<0.05.
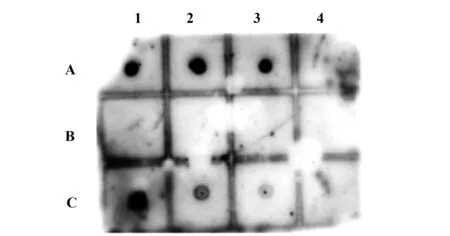
图3 斑点免疫印迹检测代表性噬菌体与rMgPa的特异结合Fig.3 Specific combination between representative phages and rMgPa was detected by Dot immunobind-ing assayNote:A1.P1;A2.P2;A3.P3;A4.P4;B1.WT;B2.BSA;B3,B4.Blank control;C1.rMgPa positive control;C2.P5;C3.P6;C4.P7.

图4 Far-western blot检测RPL35和rMgPa的相互作用Fig.4 Interaction between RPL35 and rMgPa was detected by Far-western blotNote:A.The membrane was incubated with cell total protein and B not.Lane 1.rMgPa;2.No rMgPa(blank control).
3 讨论
Ueno等[11]发现 Mg能侵入宫颈癌上皮细胞(HeLa细胞)和非癌性子宫内膜细胞(EM42细胞),但对其侵入宿主细胞后具体的致病机制研究甚少。MgPa是Mg黏附或侵入宿主细胞从而发挥其对宿主细胞致病的关键,本研究以前期表达纯化的rMgPa为靶分子,从SV-HUE-1 T7噬菌体展示cDNA文库中筛选到能与MgPa相互作用的蛋白,通过DNA序列测定和同源性分析,结果表明成功筛选到7种能与rMgPa特异结合的代表性噬菌体,通过ELISA和斑点免疫等实验表明这些代表性噬菌体能与rMgPa特异结合;Far-western blot实验进一步证实来自SV-HUC-1细胞总蛋白中的60S核糖体蛋白L35(RPL35)能与rMgPa相互作用,说明RPL35可能是rMgPa的互作蛋白。
噬菌体展示技术是当前用于研究蛋白质-蛋白质相互作用的一种快速、简便且高效的手段。相比于M13等其他噬菌体,T7噬菌体在各种极端环境(高温、低pH值)中稳定性较高,且能识别靶细胞的天然构象[12]。现今,噬菌体展示技术应用比较广泛,有学者利用噬菌体7肽库通过差减筛选得到多肽NPMIRRQ,结果证实其能与卵巢癌细胞株HO-8910细胞表面特异结合,为早期诊断和监测卵巢癌的转移复发提供了实验基础[13]。此外,T7噬菌体展示技术还可应用于模拟表位的筛选,已有研究筛选到了MUC1的两个模拟表位:AAPDFRP和SAPDDRP,并成功用于肺癌的早期诊断[14]。罗俊茜等[15]利用噬菌体展示技术筛选到能应用于膀胱癌早期诊断的小分子多肽NYZL1。潘超等[16]从噬菌体随机十二肽库中筛选到与三水白虎汤作用的类风湿关节炎滑膜成纤维细胞短肽SGVYKVAYD WQH。本课题组前期利用噬菌体展示技术筛选到能与MgPa特异结合的3条多肽,且这3条多肽能抑制Mg对SV-HUC-1的黏附[17],这些为研究MgPa蛋白与宿主细胞结合的特异作用位点提供了实验依据。
核糖体是蛋白质合成的主要场所,而核糖体蛋白是组成核糖体的主要成分,在细胞内蛋白质的生物合成中发挥重要作用。60S核糖体蛋白L35(RPL35)是核糖体亚基的一个重要组成部分,在蛋白翻译和内质网对接中发挥着重要的作用[18]。除此之外,核糖体具有参与DNA修复、细胞生长周期的调控和细胞分化等核糖体外功能,研究表明核糖体蛋白对线粒体蛋白的翻译也有一定的调控作用[19]。Amsterdam等[20]研究表明在胃癌、结直肠癌、肝癌和食管癌等肿瘤组织中会出现核糖体蛋白基因异常表达,这暗示着核糖体蛋白与肿瘤的发生息息相关。
本研究的结果说明细胞内的RPL35可能是MgPa的互作蛋白,因此,推测Mg经黏附并侵入尿道上皮细胞后,经MgPa和RPL35结合从而阻止上皮细胞蛋白质的翻译而导致细胞死亡,从而发挥其对宿主上皮细胞的致病性。下一步本课题组将研究MgPa与RPL35互作后对细胞功能的影响,从而加深对MgPa的功能理解,为进一步了解Mg的致病机制及其防治提供实验依据。
参考文献:
[1] Fernández-Molina C,Rodríguez-Preval N, Rodríguez-González I,etal.Diagnosis of Mycoplasma genitalium by MgPa and rRNA 16S gene amplification[J].Salud Publica Mex,2008,50(5):358-61.
[2] Hakre S,Casimier RO, Danboise BA,etal.Enhanced sexually transmitted infection screening for mycoplasma genitalium in human immunodeficiency virus-infected US air force personnel[J].Clin Infect Dis,2017,65(9):1585-1588.
[3] Das K,De la Garza G,Siwak EB,etal.Mycoplasma genitalium promotes epithelial crossing and peripheral blood mononuclear cell infection by HIV-1[J].Int J Infect Dis,2014, 23:31-38.
[4] Svenstrup HF,Fedder J,Abraham-Peskir J,etal.Mycoplasma genitalium attaches to human spermatozoa[J].Hum Reprod,2003,18(10):2103-9.
[5] You X, Wu Y,Zeng Y,etal.Mycoplasma genitalium-derived lipid-associated membrane proteins induce activation of MAPKs,NF-kappaB and AP-1 in THP-1 cells[J].FEMS Immunol Med Microbiol,2008,52(2):228-236.
[6] 曾焱华,吴移谋,游晓星,等.生殖支原体黏附素蛋白MgPa的表达及其多克隆抗体的制备与纯化[J].中国免疫学杂志,2010,26(11):1011-1015.
Zeng YH,Wu YM,You XX,etal.Expression of the adhesion protein of Mycoplasma genitalium and preparation and purification of it′s polyclonal antibodies[J].Chin J Immunol,2010,26(11):1011-1015.
[7] 朱有葱,何 军,曾焱华,等.人尿道上皮细胞T7噬菌体展示cDNA文库的构建与鉴定[J].生物技术,2015,25(2):151-154.
Zhu YC,He J,Zeng YH,etal.Construction and identification of T7 phage display cDNA library of SV-HUC-1cells[J].Biotechnology,2015,25(2):151-154.
[8] Sakamoto K,Ishibashi Y,Adachi R,etal.Identification of cytidine-5-triphosphate synthase1-selective inhibitory peptide from random peptide library displayed on T7 phage[J].Peptides,2017,94:56-63.
[9] Takami M,Takakusagi Y,Kuramochi K,etal.A screening of a library of T7 phage-displayed peptide identifies E2F-4 as an etoposide-binding protein[J].Molecules,2011,16(5):4278-4294.
[10] Garcia BL,Zhi H,Wager B,etal.Borrelia burgdorferi BBK32 inhibits the classical pathway by blocking activation of the C1 complement complex[J].PLoS Pathog,2016,12(1):e1005404.
[11] Ueno PM,Timenetsky J,Centonze VE,etal.Interaction of Mycoplasma genitalium with host cells:evidence for nuclear localization[J].Microbiology,2008,154(Pt 10):3033-3041.
[12] Piggott AM, Karuso P.Identifying the cellular targets of natural products using T7 phage display[J].Nat Prod Rep,2016,33(5):626-636.
[13] 王乐丹,李文桔,胡 越,等.利用噬菌体多肽库筛选卵巢癌细胞特异性结合肽[J].中国性科学,2016,25(1):56-60.
Wang YD,Li WJ,Hu Y,etal.Screening specific peptides on the surface of ovarian cancer cell by phage display[J].Chin J Hum Sexuality,2016,25(1):56-60.
[14] 邱 宇,曹 丽, 郝晓宁,等.肺癌T7噬菌体文库中MUC1蛋白抗原模拟表位的筛选[J].免疫学杂志,2016, 32(2):132-136.
Qiu Yu,Cao L,Hao XL,etal.Screening of MUC1 mimotope in T7 phage library of lung cancer[J].Immunol J,2016,32(2):132-136.
[15] 罗俊茜,张 帆,杨晓峰,等.利用体内噬菌体展示技术筛选膀胱癌特异性结合肽[J].中国免疫学杂志,2015,31(4):509-513.
Luo JX,Zhang F,Yang XF,etal.Selection of peptide specifically binding to bladder carcinoma by using phage display in vivo[J].Chin J Immunol,2015,31(4):509-513.
[16] 潘 超,肖长虹,高 燕,等.噬菌体随机十二肽库淘选三水白虎汤作用于类风湿关节炎滑膜细胞的靶点研究[J].中国免疫学杂志,2014,30(4):446-448.
Pan C,Xiao CH,Gao Y,etal.Screening target of Sanshui Baihu decoction affecting on rheumatoid arthritis synovial fibroblasts by phage display 12 peptide library[J].Chin J Immunol,2014,30(4):446-448.
[17] 朱有葱,游晓拢,邓湘赢,等.生殖支原体MgPa特异性结合多肽的筛选、鉴定与人尿道上皮细胞cDNA文库的构建[J].中华微生物学和免疫学杂志,2015,35(8):606-610.
Zhu YC,You XL,Deng XY,etal.Screening and identification of MgPa specific binding peptides and construction of T7 phage display cDNA library of SV-HUC-1 cells[J].Chin J Microbiol Immunol,2015,35(8):606-610.
[18] Jiang N,Hu L,Liu C,etal.60S ribosomal protein L35 regulates β-casein translational elongation and secretion in bovine mammary epithelial cells[J].Arch Biochem Biophys ,2015,583:130-139.
[19] Segev N, Gerst JE.Specialized ribosomes and specific ribosomal protein paralogs control translation of mitochondrial proteins [J].J Cell Biol,2018,217(1):117-126.
[20] Amsterdam A, Sadler KC,Lai K,etal.Many ribosomal protein genes are cancer genes in zebrafish [J].PLoS Biol,2004,2(5):E139.