京津风沙源治理工程不同恢复措施对草地恢复过程的差异性影响
2018-05-22李愈哲樊江文于海玲
李愈哲, 樊江文*, 于海玲
(1.中国科学院地理科学与资源研究所陆地表层格局与模拟重点实验室,北京100101;2.华侨大学化工学院,福建 厦门 361021)
草地是世界最广布的植被类型之一,面积约占全球陆地总面积的1/5[1]。中国的草原面积为3.55×108hm2,约占国土面积的41.7%,是世界草地总面积的6%~8%,居世界第二[2-3]。受人类活动和气候变化的共同胁迫,中国北方草地在过去的30年间快速退化,产生草地生产力退化、土地沙漠化、土壤流失和沙尘暴等一系列生态、环境问题[4-5]。研究显示近年来内蒙古草地退化最为明显区域主要分布在锡林郭勒草原的西部地区[4]。这一快速退化的脆弱区域位于北京西北方向约600 km,是影响北京、天津等中国核心城市群沙尘天气的最主要沙源地。为建设有效的生态屏障,中国政府发起并实施了京津风沙源治理工程、三北防护林工程和退耕还林还草工程等系列生态恢复工程,以改善区域环境条件恢复区域生态。
截至目前,全国生态工程总投资达数万亿,其中,三北、退耕还林还草、三江源、京津风沙源治理工程等全国性重大生态工程已投入4078亿元[6]。掌握重大生态工程实施的恢复效果及存在问题,以便科学优化生态工程实施方案、保障工程实施效果,使后续生态工程部署更具科学性和针对性,不仅是国家的重大迫切需求,也将促进恢复生态学研究实践和经验总结。锡林郭勒草原位于内蒙古自治区中部,地带性植被类型为典型温性草原,该草原类型约占到内蒙古天然草地总面积的1/3[7]。草原是自然环境严苛、生态系统稳定性相对脆弱的典型区域之一,不合理的人为活动已使该区50%左右的草原处于不同程度的退化之中[8]。当地自2002年开始实施京津风沙源治理工程,是工程恢复时间较长,实施的工程恢复措施及其组合最为多样的地区。因此适合作为典型区域,深入比较分析不同恢复措施对生态系统的差异性影响。
21世纪以来,国内外对生态工程效果实施后评价及恢复措施优选提升的重视程度不断上升。在国际上,美国农业部2002年发起生态保护效果评估计划(Conservation Effects Assessment Project, CEAP),分为国家尺度和区域尺度对包括湿地保护区计划、野生动物栖息地激励计划在内的7大生态保护工程采用工程区内外、工程前后和有无恢复措施对比等手段,进行工程效果评价,使用的技术方法多样全面[9],其评估理念、方法在世界范围内产生了较大的影响。如在具体工程实施流域,Brinson等[10]、Fennessy等[11]采用工程区内外对比的方式量化湿地保护计划对不同流域生态服务的提升。在工程区内多种恢复措施独自或组合实施的区域,采用量化不同恢复措施及其组合的效果再相互对比的方式,优选出刈割留茬超过30 cm对艾草榛鸡(Centrocercusurophasianus)的种群恢复效果最为明显[12-13]。此外,国际林业研究中心(CIFOR)亦基于工程区内外对比,对15个国家的森林保护项目REDD+的实施效果进行了对比,评估了工程实施对生态系统碳蓄积的作用和贡献[14]。欧盟实施的沙漠化防治评估计划(PRACTICE),则采用参与式评价的方式,对12个国家防沙治沙具体措施的有效性进行了分级评价[15]。尽管存在多样的对比方式和技术手段,工程区内外对比法有着易于操作实现、排除了气候变化等自然因素影响等优势,是最为常见和通用的量化工程实施效果的办法。
国内已有研究分析过区域性生态工程的补偿投入与产生的生态服务价值[16],这类研究主要通过计算生态服务物质量增量,进而换算价值量以评估工程的合理性。物种多样性提高对应的经济价值增长因为难以量化为货币,往往不被列入计算。生产力的变化因为缺乏地面调查数据及受所应用模型结构的限制,多不能精确描述功能群组成改变、地上地下部分比例变化等生产力精细组分的演变[5]。然而,物种多样性和生产力的组成及比例,往往受到人类土地利用方式和草地管理措施的极大影响[17-18],群落中生产力组成和多样性的改变也极大影响生态系统功能[19-20]和提供的服务[21-22]。更为重要的是,国内以往研究评估工程效益往往只着眼工程区范围内外、工程前后的改变,对具体恢复措施及其组合的差别性影响关注较少,因而评估的精细程度存在一定的不足。中国北方草原区域的退化草地恢复生态工程实施多年,应用措施丰富多样,其现存状态和恢复效果受到广泛地关注。深入解析不同恢复措施恢复效果的差异将有助于准确优选出区域内更为有效的恢复措施。总结归纳生态工程实施的区域差异性、有效性和适宜性,可以帮助后续实施工程提升管理水平和投入成效。
本研究选取京津风沙源治理工程的草原治理区作为研究区域,采用野外调查、取样测定等群落学调查方法。选取工程治理中广泛应用的恢复措施及其组合,包括围封禁牧、补播草种、小流域综合治理和人工种草4种典型工程恢复方式,对其工程区内外群落物种及生物量构成,群落物种多样性进行调查和比较。通过量化分析不同恢复措施的恢复效果异同,为优选更为高效、适宜的恢复措施,促进未来生态工程科学部署提供一定参考。
1 材料与方法
1.1 研究区域概况
京津风沙源治理工程区西起内蒙古达茂旗,东至内蒙古阿鲁科尔沁旗,南起山西代县,北至内蒙古东乌珠穆沁旗,涉及北京、天津、河北、山西及内蒙古五省(区、市)的75个县(旗)[5]。区域范围N 38°50′-46°40′,E 109°30′-119°20′,工程区域面积约4.58×105km2(图1)。工程区域广阔,覆盖了平原、山地和草地等多种不同地貌地形。因实施区域和针对问题的差异,恢复措施亦涵盖林地建设、草地恢复、水利建设和生态移民等多类项目内容和措施。本研究主要关注草原区域不同草地恢复措施对草地生态系统的影响。京津风沙源治理工程草地治理面积中,2000-2009年间,实施草地工程恢复面积共计374.6万hm2,其中围栏封育禁牧面积最大,达到295.8万hm2,其次为人工草地(36.7万hm2),基本草场建设(23.8万hm2),补播牧草(15.6万hm2)[23-24],此外在全工程区实施的水源保护措施组合小流域综合治理234.45万hm2[25],其中相当一部分实施面积分布于所调查的草原区域。本研究于2013年实施的样带调查中,主要系统调查了锡林郭勒草原区域10个工程实施旗(县)内包括围封禁牧(fenced & grazing exclusion, FG)、补播草种(replenishment seed, RS)、小流域综合治理(small watershed conservation, SW)和基本草牧场建设(basic ranch, BR)4种恢复措施对退化草地恢复的影响。
研究区域海拔1000~1400 m,属内蒙古高原中部,中纬度西风带半干旱,干旱季风性气候。年均气温-0.4 ℃,1月和7月平均气温分别为-23 ℃和17.9 ℃,年均降水量350 mm,主要集中在6-8月,全年以大风干燥天气为主[26]。土壤类型以栗钙土为主[27]。
当地主要地带性气候顶级植被为典型温性草原,如大针茅群落(Form.Stipagrandis)、克氏针茅群落(Form.Stipasareptanavar.krylovii)等,因地形差异及草地退化局部亦存有区域性草甸草原及荒漠化草原。自然群落主要建群植物为大针茅(Stipagrandis)、羊草(Leymuschinensis)、糙隐子草(Cleistogenessquarrosa)、克氏针茅(Stipasareptanavar.krylovii)、冷蒿(Artemisiafrigida)等,常见优势种还包括小叶锦鸡儿(Caraganamicrophylla)、羽茅(Achnatherumsibiricum)、冰草(Agropyroncristatum)、米氏冰草(Agropyronmichnoi)、芨芨草(Achnatherumsplendens)等。
1.2 研究方法
1.2.1样地设置 野外调查于2013年8月(全年群落生物量最大)开展。依据内蒙古农业大学提供的各旗(县)工程实施区域分布图,结合调查区域附近牧民的问卷调查结果,本研究在京津风沙源治理工程实施范围内的10个旗(县)选取设置了28个恢复措施及对照观测点(图1)。每个地点设置3个生物量调查样地,以及 10个随机抛框用于记录测定群落物种组成、群落物种频度。每个生物量调查样地内设置3个生物量调查刈割样方,用于调查群落结构及生物量组成。28个观测点中包含4种具有代表性的工程恢复措施,其中围封禁牧站点6个,补播草种的观测点3个,小流域综合治理观测点3个,基本草牧场建设观测点2个。本研究在每个工程观测点围栏外毗邻地形环境一致的自由放牧区域(距离恢复措施围栏最近约400 m左右)都设置有一个对照观测点用来配对比较,共计14个对照观测点以量化准确评估恢复措施对草地生态系统的改变和影响。

图1 研究区与采样点分布Fig.1 Study area and survey site distribution
所有恢复措施站点位置在空间上相互远离交错,避免个别工程实施区域受到区域特殊地形、小气候等自然因素及个案性管护条件带来的影响,从而减少因取样点选择造成恢复措施效果评价偏差。
1.2.2恢复措施 研究所选取的恢复措施中,围封禁牧措施是通过围栏设施阻止牛羊等家养牲畜进入草地,完全排除了牲畜对草地的践踏和采食影响,以恢复草地状态和功能的一种工程恢复措施。在本研究中,各观测点的禁牧时间有所不同,一般为3~8年。补播草种措施是在春季人工补充播撒牧草种子,依据退化程度及区域差别,补播的牧草种类有所差别。在京津风沙源治理工程本研究调查涉及实施区,补播的牧草种类主要包括羊草、垂穗披碱草(Elymusnutans)等饲用价值较高的当地天然牧草。本研究中调查的基本草牧场建设区域实施的恢复措施为翻耕后人工种植牧草。京津风沙源治理工程中小流域治理是以30 km2以上,最大不超过50 km2的小流域为单元,合理确定流域内每一个地块的土地利用方向及水土保持技术措施,做到互相协调,互相配合,形成综合的以防治水土流失为主要目标的措施体系。其具体措施包括打坝淤地、坡面截流沟、补播灌草恢复植被、拦沙坝、草方格等。
1.2.3群落特征和物种组成 每个调查地点(包括各恢复措施地点和对照点)随机抛取频度调查样方(1 m×1 m)10个,仅记录所有抛框内植物物种出现次数。同时记录群落地形、地貌特征和土壤质地,GPS定位样地经纬度和海拔。每个调查地点选取3个地形条件一致,同时能较好代表群落整体状态的25 m×25 m样地,按照对角线取样法在其中设置3个1 m×1 m的样方,采用草地群落学调查方法,测定记录样方内所有物种组成,群落最大高度、平均高度,群落总盖度,并分种测量最大高度、平均高度,估计各物种分盖度。
1.2.4地上及地下生物量测定 利用收获法测定样方地上生物量,在每个样地的3个1 m×1 m样方重复里齐地面刈割样方内植物,并分种放入信封,在烘箱中恒温65 ℃烘干48 h至恒重后称重记录[28]。地下生物量的测定采用土钻法(直径70 mm)[29-31],在地上部分刈割完毕的每个样方内,按照对角线取土钻3钻,每钻分3层各取10 cm土柱(0~10 cm, 10~20 cm, 20~30 cm)。同样方同层的3份钻样混合作为一个地下生物量样品放入自封袋,即每样地类型分3层取样,每层样品重复为3个。样品放入0.3 mm的网袋,水洗分离土壤和根系,随后用0.3 mm网筛分离石砾等非根杂质,在65 ℃的烘箱内烘干48 h至恒重并称重。
1.2.5物种多样性测定与计算方法 1)物种丰富度:
S=调查中出现在样地内的物种总数
2)多样性指数
Gleason指数:dGl=S/lnA
Shannon-Wiener指数:H′=-∑PilnPi
3)均匀度指数
Pielou指数:J=H′/lnS[32]
式中:S为物种种数;A为样方面积(m2);Pi为种i的重要值,重要值(IV)的计算公式[33]为:
IV=(相对高度+相对盖度+相对频度)/3
1.2.6工程恢复方式影响计算方法 因样点设置采用了内外对比法。根据配对的工程恢复站点及其对应对照站点的状态数据和相关计算指数,根据下式估算工程恢复措施实施对群落原有状态的影响程度。
工程恢复影响:I=(Ip-Ick)/Ick×100%
式中:I为测算的工程恢复措施实施造成的影响程度;Ip为工程恢复区域内站点生态系统的测定状态及相关计算指数;Ick为对应的对照站点的生态系统的测定状态及相关计算指数。
1.3 数据分析
使用Excel进行数据初步整理,采用Origin 8.0作图,采用SPSS 12.0进行数据统计分析。不同恢复措施及对照组间植被特征、生物量及其组成和物种多样性特征差异使用单因素方差分析(one-way ANOVA)进行比较,采用Turkey检验。各工程恢复措施与其对照间差异采用配对T检验(PairedT-test)。不同工程恢复方式对生物量及其组成和多样性指标的影响差异采用单因素方差分析进行比较,采用Turkey检验。
2 结果与分析
2.1 恢复措施对群落结构特征与物种组成的影响
本研究将调查的28个站点,依据工程实施状态和实施恢复措施归纳为4种工程恢复方式及相应的4组对照共计8类群落(表1)。分析结果表明,与各自对照相比4种工程恢复方式都趋向增加群落盖度,其中围封禁牧的盖度增加达到显著水平(表1,P<0.05),增幅约50%。4种工程恢复方式都趋向增加群落高度,其中围封禁牧、补播草种和基本草牧建设对群落高度的增加达到显著水平,在增加最为明显的基本草牧场建设区域内,群落高度平均增长3.1倍。各站点的建群种和优势种(群落中3个IV最大的物种)被列于表1中,同组不同站点间重复物种进行了归并。
2.2 恢复措施对群落生物量的影响
2.2.1群落地上生物量的构成和差异 与各自对照组相比,各工程恢复方式均趋向增加群落的地上生物量(图2),其中长期围封禁牧、草种补播和基本草牧场建设的增加达到显著水平(P<0.05)。在4种不同恢复措施中,基本草牧场建设的地上生物量最高,平均为690.0 g·m-2, 显著高于其他恢复方式。相比其对照,生物量增幅为235%,远高于其他3种恢复方式。另外3种恢复方式(围封禁牧、草种补播和小流域治理)地上生物量均在200.0 g·m-2左右,彼此间差异不显著。4种不同恢复方式的对照组之间亦无显著差异。
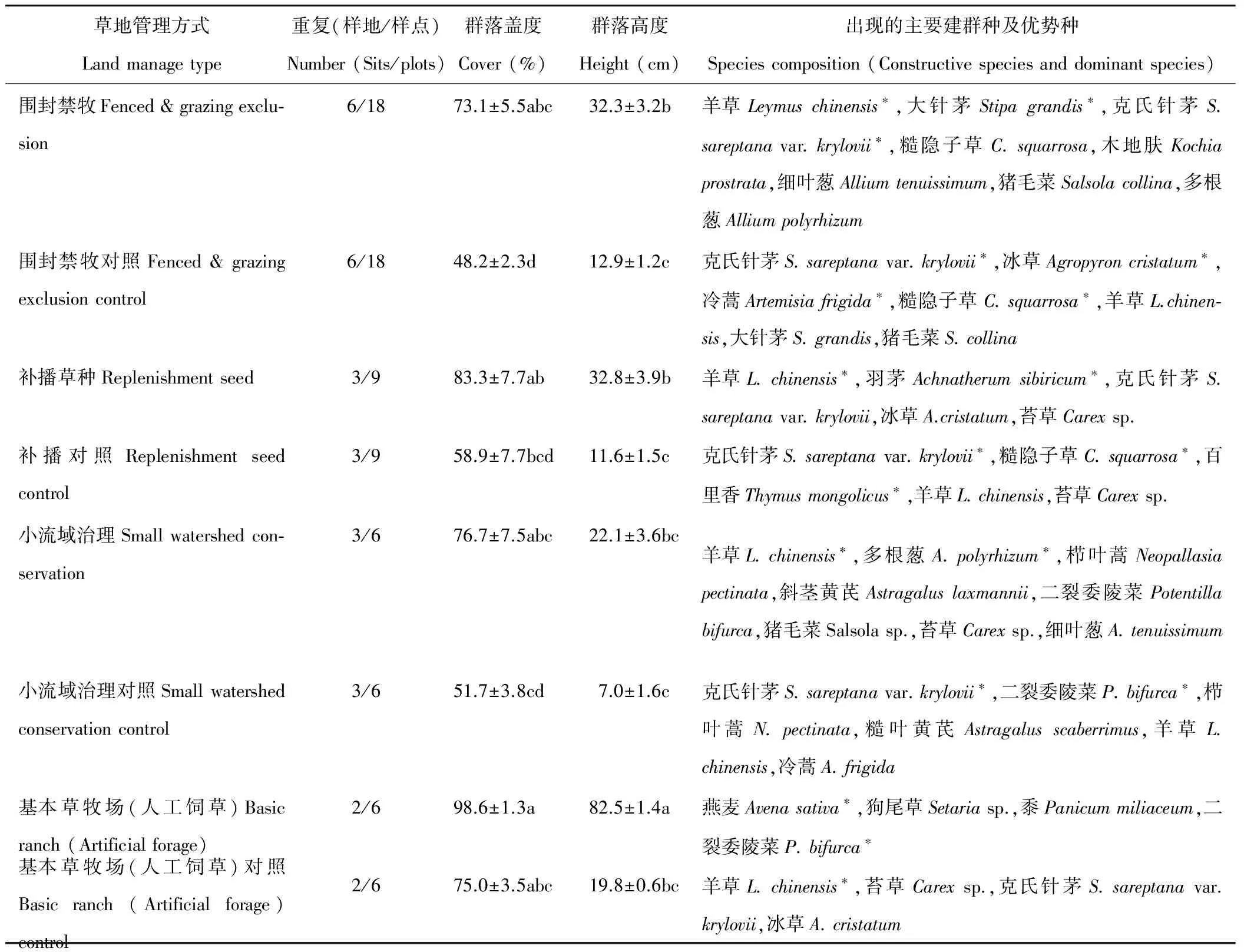
表 1 不同管理方式下群落特征及物种组成Table 1 Community characteristics and species composition in different grassland manage types
在植物类群功能群方面,不同的工程恢复方式都趋向增加群落中的禾本科植物比例,同时降低群落的杂类草比例(图3a)。其中补播草种区域与对照相比,杂类草占比下降了82%。而莎草科、豆科和菊科植物的地上生物量比例没有明显的规律性变化。与对照群落相比,不同工程恢复方式还趋于增加群落中的C3植物比例(图3b)。其中围封禁牧和补播草种对C3植物比例的增加更为可观,相比各自对照,前者增加40%,后者增加29%。小流域治理和基本草牧场建设对群落C3植物比例的增加幅度很小。
2.2.2群落地下生物量的组成和差异 各群落的地下部分生物量均大幅高于地上生物量,基本草牧场建设区域的地下生物量较低,平均为1500 g·m-2左右,而最高的补播草种恢复区域群落地下生物量平均达3200 g·m-2左右,约为前者的两倍。不同的工程恢复方式和具体管理措施对群落地下生物量的影响呈现极大的差异性。其中,围封禁牧能显著提高群落的地下生物量(图4,P<0.05),而基本草牧场建设则显著降低群落的地下生物量(P<0.05)。补播草种趋向于增加群落的地下生物量,但增加未达显著水平(P>0.05)。小流域治理与其对照组的地下生物量彼此间差异很小(P>0.05),恢复措施对群落地下生物量几乎无改变。

图2 不同工程恢复方式及其对照的总地上生物量Fig.2 Aboveground biomass in different managepatterns and their controls
2.2.3群落的总生物量和地上地下比 各工程恢复区域中,平均总生物量最高的是补播草种区域,为2966 g·m-2,最低的是基本草牧场建设区域,平均为2156 g·m-2,4种工程恢复方式间没有显著差异。各对照区域中,最高的是基本草牧场对照区域,总生物量平均为2979 g·m-2,显著高于最低的围封禁牧对照区域(图5,P<0.05),而与其余两工程方式的对照区域差异不显著(P>0.05)。相比各自的对照区域,围封禁牧显著增加群落的总生物量(P<0.05),小流域治理和补播草种趋向于增加群落的总生物量,但增加不显著(P>0.05)。基本草牧场建设区域的总生物量趋向于减少,变化未达显著水平(P>0.05)。除基本草牧场建设区域外,群落的地上、地下部分比例彼此间差异不大,各工程恢复方式多趋向于增加地上部分所占比例,降低地下部分所占比例。除基本草牧场外,各组群落的地下部分比例均高于90%,而地上部分低于10%。在基本草牧场建设区域,群落的地上部分比例显著增加至(32.3±2.5)%,而地下部分比例则下降至(67.8±2.5)%(图5,P<0.05)。

图3 不同工程恢复方式及其对照的地上生物量植物功能群组成Fig.3 The plant functional group compositions of aboveground biomass in different manage patterns of the conservation project and their controls
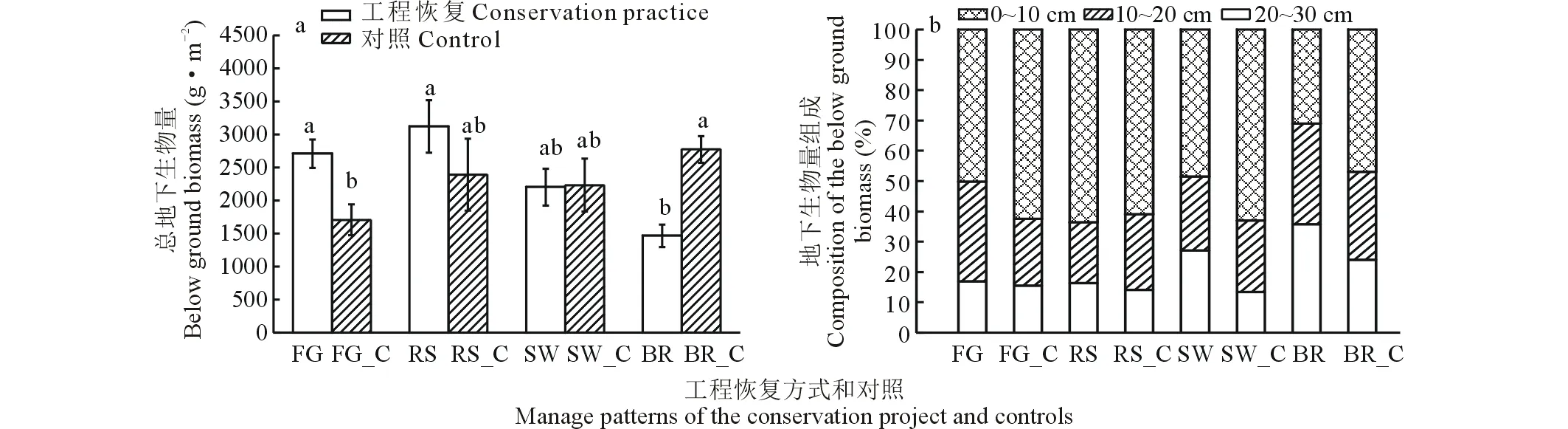
图4 不同工程恢复方式及其对照群落地下生物量及其分层构成的影响Fig.4 The below ground biomass in different manage patterns also their controls of the conservation project and the composition of below ground biomass in three depth layers

图5 不同工程恢复方式及其对照的群落总生物量及地上、地下生物量所占比例Fig.5 Total biomass in different manage patterns also their controls of the conservation project and the percentage of above/below ground biomass of communities
2.3 恢复措施对物种多样性的影响
物种丰富度是群落的重要特征[34],在各组样地中,围封禁牧物种丰富度最高,而基本草牧场对照群落物种丰富度最低。不同工程恢复方式相比各自的对照群落,除补播外,其他3种恢复措施都趋向于增加群落的物种丰富度,其中基本草牧场建设区域的增加达到显著水平(表2,P<0.05)。
物种多样性和均匀度水平一定程度反映了群落的稳定性和发育阶段。本研究测算的两种物种多样性指数(Gleason指数和Shannon-Wiener指数):dGl在各组群落间变化情形与物种丰富度相近,但变化并不显著(P>0.05)。而H′显示,各工程恢复措施的应用均趋向于降低群落的物种多样性,其中在补播草种和基本草牧场建设区域的下降达到显著水平(表2,P<0.05)。不同工程恢复方式与其各自对照组相比,群落的均匀度均趋向于下降,其中补播草种和基本草牧场建设下降最为明显(P<0.05),而围封禁牧对群落均匀度的影响最小(P>0.05)。
2.4 工程恢复方式对群落的差异性影响
各工程恢复方式均有助于群落的地上生物量恢复,其中围封禁牧、补播草种和基本草牧场建设区域的地上生物量均显著增加。且基本草牧场建设和围封禁牧对群落地上生物量的增加显著高于其他两种恢复方式(图6,P<0.05)。不同工程恢复方式对群落地下生物量的影响存在明显差异,其中围封禁牧和补播牧场均趋向增加群落的地下生物量,围封禁牧管理方式对地下生物量的增加达到显著水平(58%,P<0.05)。小流域治理和基本草牧场建设趋向降低群落的地下生物量,其中基本草牧场建设对群落地下生物量的降低达到显著水平(49%,P<0.05)。不同工程恢复方式对群落总生物量的影响情形与地下生物量更为接近。围封禁牧显著增加群落总生物量(87%,P<0.05),基本草牧场建设显著降低群落总生物量(29%,P<0.05),补播草种和小流域治理增加群落的总生物量,但不显著(P>0.05)。

表2 不同工程恢复方式及其对照的群落物种多样性特征Table 2 Community species diversity in different manage patterns also their controls of the conservation project

图6 不同工程恢复方式对群落总生物量、地上和地下生物量的影响差异Fig.6 Comparison of different restoration practices effect on community total biomass, above and below ground biomass
基本草牧场建设显著增加群落的物种丰富度(图7,P<0.05),而补播草种则显著降低群落的物种丰富度(P<0.05),其余两种工程恢复方式趋向增加群落物种丰富度,但工程方式之间对物种丰富度影响差异并不显著(P>0.05)。各工程恢复方式对群落Gleason指数的影响均未达显著水平,恢复方式之间影响无显著差别(P>0.05)。不同工程恢复方式对群落Shannon-Wiener指数和Pielou均匀度指数的影响相似,都趋向于增加。其中Shannon-Wiener指数和Pielou均匀度指数在基本草牧场建设和补播草种措施区域的减少达到了显著水平。补播草种对群落H′影响最为明显,平均下降35%,影响显著高于小流域治理和围封禁牧。基本草牧场建设对物种多样性的抑制也明显强于围封禁牧(P<0.05)。基本草牧场建设对J影响最为明显,平均下降37%,与小流域治理和围封禁牧产生的影响存在明显差异(P<0.05)。
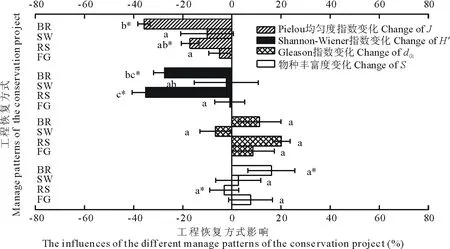
图7 不同工程恢复方式对群落植物物种多样性特征的影响差异Fig.7 Comparison of different restoration practices effect on the plant community species diversity indices
3 讨论
3.1 工程恢复措施实施对草地群落特征、物种组成的影响
草地利用和管理方式的转变常直接影响地表的植被覆盖状况和群落的组成[35]。工程恢复措施作为一类以恢复为目的的管理方式,其造成的地表植被的改变往往具体表现为群落物种结构组成、生物多样性水平和生产力水平变化等群落改变[36]。本研究中,4种不同工程恢复措施均不同程度增加了草地植被盖度(30%~52%),其中围封禁牧的增加达到了显著水平,增加幅度达到52%(表1)。这与Wu 等[5]利用遥感方法观测到的增加趋势相吻合,尽管比其结果11.8%增加幅度更高,这一差异应归因于遥感研究中不能准确地区分识别工程和非工程实施区,而工程区域的植被恢复才是全区域植被覆盖增加的主要贡献者。以往研究将工程恢复区植被生产力的增长80%归因人类活动,特别是大规模生态工程的恢复作用[37]。与其结论相应证,本研究观察到各工程恢复方式明显地增加了植被的群落高度,在4种恢复方式中除小流域治理外,其他3种方式的植被高度增加均达到显著水平(P<0.05),冠层高度大致是放牧对照区域的3倍(表1)。牲畜采食减少致使的生物量积累是植被平均高度增加的最主要原因。草地植被高度和盖度的增加无疑有效加强了植被生物量的增长和累积,也将帮助地表土壤保持对抗风蚀水蚀,还帮助区域植被碳库的建立、恢复,通过对碳循环的影响有助于缓解气候变化。
围封禁牧、补播牧草和小流域治理这3种恢复方式与其对照区域相比,如表1所示,群落建群种由冰草、冷蒿、糙隐子草、百里香、委陵菜等较为低矮的植物转变为羊草、大针茅、羽茅等较高大的多年生草本植物。甚至相比大针茅更为低矮[38]的克氏针茅的群落重要值在补播草种措施的影响下也明显下降。这一现象可能由两方面原因共同驱动。羊草的适口性和营养价值高于百里香、委陵菜和冷蒿等有特殊气味的杂类草[39],对照区域的采食强度相比工程恢复区更大,因此采食减少有助于这些优良牧草物种的增加和恢复。此外,因为恢复措施对植被高度和盖度的恢复,更为高大的羊草、羽毛和大针茅相比低矮物种具有更强的光竞争优势[17],其高强度的遮盖效应会抑制和限制低矮植物的生长[17,19],逐渐取代其建群种地位。群落中重新确立及重要值大幅增加的建群种都是多年生较高禾本科植物,其更为强大和多年保持的根系有助于降低区域土壤的可蚀性,土壤中有机物的更多积累还将改善生态系统的土壤团粒结构,正反馈于植物的生长、沙尘的固定。更高建群种的确立还会通过消光系数的调节影响群落内光照条件和能量分配,尤其是地下层植物可获取的光照大幅减少,将对植物群落物种组成和多样性产生系列后果。
桥本氏甲状腺病是一种自身免疫性甲状腺疾病,临床上可以按照甲状腺功能水平,分为甲亢期、甲减期和恢复期,本文所讨论的是,桥本氏甲状腺病甲减期病例,药物治疗是这类疾病有效的治疗方式[1] 。而除了常规的西药治疗外,中药治疗的加入,有利于更好的改善桥本氏甲状腺功能减退患者的临床症状,本文就两种不同的用药方式所取得的效果,进行讨论分析。研究样本群体选自我院的42例患者,以下为详细研究过程汇报。
3.2 不同工程恢复措施对草地生产力的差异性影响
尽管已有研究通常认为中国生态工程的实施增加了区域的生产力[37]和生物量[5],这与国外实施的系列生态工程效果评估结果相似[10,14]。但这些研究大都只反映了区域植被地上部分的明显恢复。然而需要注意到,植物地上部分即地上生物量增加,不能反映其群落物种构成变化造成的生物量组成质量的改变,也不体现占比约90%的草原地下生物量的变化情形[18]。
本研究调查结果显示,在植物的科属组成上,最一致而明显的改变是各工程恢复方式均提升了群落中禾本科植物生物量占比,莎草科植物比例趋向于小比例增长,而豆科、菊科和杂类草生物量占比趋向于下降(图3a)。这反映了亚欧典型温性草原地带性顶级的建群种主要是针茅属或其他高大禾本科植物,恢复措施使建群种重新确立及重要值上升的过程,必然增加群落中禾本科植物生物量的比重。而草原中豆科、菊科和杂类草一般较前者低矮,在光竞争中处于劣势,在群落中比重因此不断下降。这一变化使得群落中可食和优良牧草的比例上升,有助于退化草地在生产性能上的恢复。在C3和C4植物构成方面,围封禁牧和补播草种均趋向于增加群落中的C3植物生物量比例(图3b),降低C4植物比例。一般认为草地中C4植物往往是水分利用效率更高,更为耐旱的物种[20]。因此恢复措施的实施有促使群落由旱生向中生演变的趋势,在演替理论中气候顶级有着群落在当地环境条件允许下可达到的最大中生性,因此这一变化符合恢复演替理论,侧面反映了群落组成朝气候顶级的恢复性演变。C3植物总量和比例上升增加群落耗水,增加的水分消耗可能贡献自植被恢复后对降水更好的截留和拦蓄作用。同时,除基本草牧场建设以外,其余3个工程恢复措施下群落中多年生草本植物比例的增加和一年生草本植物的下降(图3c)也证实了群落组成的恢复性演替过程。而基本草牧场建设中多年生植物被一年生植物取代完全是人工翻耕种植一年生饲草的结果,不具普遍意义。
只有掌握群落地下生物量对不同恢复措施的响应[3],才能准确把握群落总生物量积累对不同恢复措施及其组合的差异性响应。调查中,尽管各恢复措施均明显增加群落的地上生物量,不同工程方式对地下生物量的影响却呈现很大分歧,补播草种和围封禁牧趋向增加地下生物量,其中围封禁牧达到显著水平,而基本草牧场建设中的人工种草措施则显著降低地下生物量(图4,6)。研究发现并非以往关注的地上生物量[5, 37],而是地下生物量在不同恢复措施影响下的差异性改变主导了群落总生物量的变化趋势。研究结果显示结合地下生物量后,只有围封禁牧、补播草种和小流域综合治理趋向于增加群落的总生物量,其中只有围封禁牧的增加(58%)达到显著水平(图5,6),而基本草牧场中的人工种草措施反而显著降低群落的总生物量(P<0.05)。这主要是由于人工种植饲草作物多为一年生植物,与其他恢复方式中植物群落相比,其损失了多年生植物长时间积累的地下根系。尽管其地上生物量在4种恢复方式增加幅度最大(超过2倍,图2),能最大程度优化草地的饲草供应功能,但其减少草地碳库,削弱生态系统碳汇功能将产生负面的环境影响。
3.3 不同工程恢复措施对植物群落物种多样性的差异性影响
以往研究认为短期的禁牧,尤其是对退化草地,不仅恢复牧草产量,还不同程度增加群落的物种多样性[18-19]。物种多样性在全球尺度上,也被广泛认为与生产力存在正相关关系。然而本研究中,尽管群落的地上生物量普遍增加(图2),但多样性却呈现出更为复杂的变化情形(表2,图7)。尽管各工程恢复方式下群落Gleason指数的变化并不一致且均不显著,但Shannon-Wiener多样性指数在不同工程恢复措施下均趋向下降,其中基本草牧场建设和补播草种恢复方式下达到了显著水平(P<0.05)。尽管生态工程能够较好的恢复地表植被覆盖和生产力(表1,图6),帮助恢复生态系统固定沙源、提供饲用牧草和调节碳汇等重要生态功能,但其对群落植物物种多样性的影响可能并不符合工程实施最初的预期。对于不同工程恢复措施普遍趋向降低草地植物群落物种多样性的现象,猜测了如下可能原因,并分析了降低过程、机制。群落中生产力增加而多样性下降的现象,在以往的氮素添加实验中被广泛报道[17,40-42],为解释这一现象而提出的光竞争假说[43]和凋落物积累假说[44]亦可部分适用于工程恢复措施造成的影响。比照光竞争理论,工程恢复措施不同程度减少了牲畜对草地植被的采食、践踏压力,工程恢复区域那些被牲畜偏好的高大优良牧草在恢复措施作用下快速恢复,在高度、盖度上占得优势,荫蔽一些耐践踏、牲畜不喜食、生长较慢或矮小的物种(表1)。恢复措施使得群落中物种原本依靠对采食和践踏的耐受和负选择获取的优势转变为在光竞争中的劣势,部分生物量较小的低矮物种可能因此消失。此外,在恢复措施影响下,随着植物生产力提高,凋落物积累量也将增加,阻止了一些种子的萌发和幼苗生长,不利于一年生植物而有利于多年生植物。这部分解释了观察到的群落功能群组成改变(图3c)。由此群落更新过程和格局的改变也可能是群落物种组成和多样性降低的可能原因。
光竞争加强将导致群落中物种重要值的极化,生物量多、高大的物种将获取更多的光资源而愈发增强其群落优势,劣势物种愈发受到抑制,群落内物种构成变得极不均衡。本研究中Pielou均匀度指数在各恢复措施群落中均趋向下降,部分措施降低幅度接近40%(P<0.05,图7)。尽管群落中植物种的种数在不同措施下并没有显著减少。但物种多样性是把物种丰富度与物种均匀度结合起来考虑的。因此在群落均匀度出现严重下降的情况下,各工程方式又不能显著增加群落中的物种种类数,H′在各恢复措施一致趋向降低就是必然的结果。而放牧的对照组草地凋落物积累量较少,采食和践踏降低了植被高度和盖度,帮助光照到达下层植被,缓解物种间光竞争,因而有助于维持群落的物种多样性。更有研究指出适度地干扰如放牧和刈割可以提高物种丰富度,改善种群的生态位配置以最大限度地利用环境资源,更符合中度干扰理论中群落达到最大物种多样性的条件。
4 结论
京津风沙源治理工程4种代表性工程恢复措施及措施组合均能不同程度恢复植被的地上生物量,有利于区域沙尘固定和草地生产功能恢复。群落物种和功能群构成整体趋向中生性顶级演变。虽然种植人工饲草的基本草牧场能最大程度增加群落的地上生物量,快速恢复草地生产功能,但群落地下生物量大幅下降,降低了群落总生物量,造成草地生物质碳库流失。工程恢复措施并不能大幅改变群落的物种丰富度,而人工饲草种植和补播草种工程措施由于大幅降低群落均匀度,能显著降低植物群落的物种多样性(Shannon-Wiener多样性指数),而长期围封禁牧措施对草地物种多样性的抑制作用相对更小。
参考文献References:
[1] Scurlock J, Hall D. The global carbon sink: a grassland perspective. Global Change Biology, 1998, 4(2): 229-233.
[2] Hu Z M, Fan J W, Zhong H P,etal. Progress on grassland underground biomass researches in China. Chinese Journal of Ecology, 2005, 24(9): 1095-1101.
胡中民, 樊江文, 钟华平, 等. 中国草地地下生物量研究进展. 生态学杂志, 2005, 24(9): 1095-1101.
[3] Fan J, Zhong H, Harris W,etal. Carbon storage in the grasslands of China based on field measurements of above-and below-ground biomass. Climatic Change, 2008, 86(3): 375-396.
[4] Zhang Q Y, Wu S H, Zhao D S,etal.Temporal-spatial changes in Inner Mongolia grassland degradation during past three decades. Agricultural Science & Technology, 2013, 4: 676-683.
张清雨, 吴绍洪, 赵东升, 等. 30 年来内蒙古草地退化时空变化研究. 农业科学与技术, 2013, 4: 676-683.
[5] Wu Z, Wu J, He B,etal. Drought offset ecological restoration program-induced increase in vegetation activity in the Beijing-Tianjin Sand Source Region, China. Environmental Science & Technology, 2014, 48(20): 12108-12117.
[6] Ouyang Z Y, Wang Q, Zhen H,etal. National remote sensing survey and assessment of ecological and environmental change (2000-2010). Beijing: Science Press, 2014.
欧阳志云, 王桥, 郑华, 等. 全国生态环境十年变化(2000-2010年)遥感调查与评估. 北京: 科学出版社, 2014.
[7] Hugejiletu,Yang J, Baoyintaogetao,etal. Effects of different disturbances on species diversity and biomass of community in the typical steppe. Acta Prataculturae Sinica, 2009, 18(3): 6-11.
呼格吉勒图, 杨劼, 宝音陶格涛, 等. 不同干扰对典型草原群落物种多样性和生物量的影响. 草业学报, 2009, 18(3): 6-11.
[8] Li B.The rangeland degradation in North China and its preventive strategy. Scientia Agricultura Sinica, 1997, 30(6): 1-9.
李博. 中国北方草地退化及其防治对策. 中国农业科学, 1997, 30(6): 1-9.
[9] Mausbach M J, Dedrick A R. The length we go. Journal of Soil and Water Conservation, 2004, 59(5): 96.
[10] Brinson M M, Eckles S D. US department of agriculture conservation program and practice effects on wetland ecosystem services: a synthesis. Ecological Applications, 2011, 21(supple 1): 116-127.
[11] Fennessy S, Craft C. Agricultural conservation practices increase wetland ecosystem services in the Glaciated Interior Plains. Ecological Applications, 2011, 21(supple 1): 49-64.
[12] Rodgers R D, Hoffman R W. Prairie grouse population response to conservation reserve program grasslands: an overview// Proceedings of the conservation reserve program national conference, planting for the future, fort collins. CO: United States Department of the Interior and United States Geological Survey, F, 2004.
[13] Schroeder M A, Vander Haegen W M. Response of greater sage-grouse to the conservation reserve program in Washington State. Studies in Avian Biology, 2011, 38: 517-529.
[14] Börner J, Baylis K, Corbera E,etal. Emerging evidence on the effectiveness of tropical forest conservation. PloS one, 2016, 11(11): e0159152.
[15] Rojo L, Bautista S, Orr B J,etal. Prevention and restoration actions to combat desertification. An integrated assessment: The PRACTICE Project. Sécheresse, 2012, 23(3): 219-226.
[16] Zheng H, Robinson B E, Liang Y C,etal. Benefits, costs, and livelihood implications of a regional payment for ecosystem service program. Proceedings of the National Academy of Sciences, 2013, 110(41): 16681-16686.
[17] Zhao J, Li W, Jin G H,etal. Responces of species diversity and aboveground biomass to nitrogen addition in fenced and grazed grassland on the Loess Plateau. Acta Prataculturae Sinica, 2017, 26(8): 54-64.
赵洁, 李伟, 井光花, 等. 黄土区封育和放牧草地物种多样性和地上生物量对氮素添加的响应. 草业学报, 2017, 26(8): 54-64.
[18] Li Y Z, Fan J W, Zhang L X,etal. The impact of different land use and management on community composition, species diversity and productivity in a typical temperate grassland. Acta Prataculturae Sinica, 2013, 22(1): 1-9.
李愈哲, 樊江文, 张良侠, 等. 不同土地利用方式对典型温性草原群落物种组成和多样性以及生产力的影响. 草业学报, 2013, 22(1): 1-9.
[19] Li Y, Fan J, Hu Z,etal. Comparison of evapotranspiration components and water-use efficiency among different land use patterns of temperate steppe in the Northern China pastoral-farming ecotone. International Journal of Biometeorology, 2016, 60(6): 827-841.
[20] Li Y, Fan J, Hu Z,etal. Influence of land use patterns on evapotranspiration and its components in a temperate grassland ecosystem. Advances in Meteorology, 2015, (2015): 1-12.
[21] Liu J, Li S, Ouyang Z,etal. Ecological and socioeconomic effects of China’s policies for ecosystem services. Proceedings of the National Academy of Sciences, 2008, 105(28): 9477-9482.
[22] Ouyang Z, Zheng H, Xiao Y,etal. Improvements in ecosystem services from investments in natural capital. Science, 2016, 352(6292): 1455-1459.
[23] Shi S, Zou X Y, Zhang C L. Investigation of vegetation restoration in the area of Beijing and Tianjin sand storm source control project. Science of Soil and Water Conservation, 2009, 7(2): 86-92.
石莎, 邹学勇, 张春来. 京津风沙源治理工程区植被恢复效果调查. 中国水土保持科学, 2009, 7(2): 86-92.
[24] Zhang L, Cao W, Fan J. Soil organic carbon dynamics in Xilingol grassland of northern China induced by the Beijing-Tianjin Sand Source Control Program. Frontiers of Earth Science, 2017, 11(2): 407-415.
[25] Li M Z, Zhang X P. Problems and countermeasures in implementing sandustorm source control project in and around Beijing and Tianjin. Journal of Beijing Forestry University (Social Sciences Edition), 2004, 3(3): 76-79.
李明志, 张学培. 实施京津风沙源治理工程的问题及对策. 北京林业大学学报(社会科学版), 2004, 3(3): 76-79.
[26] Zhao X Y, Wang S P. Responses of the anatomical characteristics of plant leaf to long-term grazing under different stocking rates in Inner Mongolia steppe. Acta Ecologica Sinica, 2009, 29(6): 2906-2918.
赵雪艳, 汪诗平. 不同放牧率对内蒙古典型草原植物叶片解剖结构的影响. 生态学报, 2009, 29(6): 2906-2918.
[27] Wang Y F, Chen Z Z, Tieszen L T. Distribution of soil organic carbon in the major grasslands of Xilinguole, Inner Mongolia, China. Acta Phytoecologica Sinica, 1998, 22(6): 545-551.
王艳芬, 陈佐忠, Tieszen L T. 人类活动对锡林郭勒地区主要草原土壤有机碳分布的影响. 植物生态学报, 1998, 22(6): 545-551.
[28] Fan J W, Du Z C, Zhong H P. Red clover and cocksfoot biomass and spatio-temporal structure of their leaf area. Acta Agrestia Sinica, 2004, 12(3): 204-208.
樊江文, 杜占池, 钟华平. 红三叶, 鸭茅生物量和叶面积时空结构特征. 草地学报, 2004, 12(3): 204-208.
[29] Yu W T, Yu Y Q. Advances in the research of underground biomass. Chinese Journal of Applied Ecology, 2001, 12(6): 927-932.
宇万太, 于永强. 植物地下生物量研究进展. 应用生态学报, 2001, 12(6): 927-932.
[30] Song L L, Fan J W, Zhong H P,etal. Changes of biomass and species richness of grassland community along an altitude gradient in Hongchiba, Chongqing. Acta Agrestia Sinica, 2010, 18(2): 160-166.
宋璐璐, 樊江文, 钟华平, 等. 重庆红池坝地区草地群落生物量及物种丰富度沿海拔梯度格局变化. 草地学报, 2010, 18(2): 160-166.
[31] Liu F S, Zhou Z B, Hu S J,etal. Influence of different soil coring methods on estimation of root distribution characteristics. Acta Prataculturae Sinica, 2012, 21(2): 294-299.
刘凤山, 周智彬, 胡顺军, 等. 根钻不同取样法对估算根系分布特征的影响. 草业学报, 2012, 21(2): 294-299.
[32] Magurran A E. Ecological diversity and its measurement. NJ: Princeton University Press Princeton, 1988.
[33] Sun J, Li X Z, Hu Y M,etal. Classification, species diversity, and species distribution gradient of perma frost wetland plant communities in Great Xing an Mountains valleys of Northeast China. Chinese Journal of Applied Ecology, 2009, 20(9): 2049-2056.
孙菊, 李秀珍, 胡远满, 等.大兴安岭沟谷冻土湿地植物群落分类, 物种多样性和物种分布梯度. 应用生态学报, 2009, 20(9): 2049-2056.
[34] Wang S P, Li Y H. Influence of different stocking rates on plant diversity ofArtemisiafrigidacommunity in Inner Mongolia steppe. Acta Botanica Sinica, 2001, 43(1): 89-96.
汪诗平, 李永宏. 不同放牧率对内蒙古冷蒿草原植物多样性的影响. 植物学报, 2001, 43(1): 89-96.
[35] Miao F H, Guo Y J, Miu P F,etal. Influence of enclosure on community characteristics of alpine meadow in the northeastern edge region of the Qinghai-Tibetan Plateau. Acta Prataculturae Sinica, 2012, 21(3): 11-16.
苗福泓, 郭雅婧, 缪鹏飞, 等. 青藏高原东北边缘地区高寒草甸群落特征对封育的响应. 草业学报, 2012, 21(3): 11-16.
[36] Li X B. A review of the international researches on land use/land cover change. Acta Geogrphica Sinica, 1996, 51(6): 553-558.
李秀彬. 全球环境变化研究的核心领域. 地理学报, 1996, 51(6): 553-558.
[37] Mu S, Zhou S, Chen Y,etal. Assessing the impact of restoration-induced land conversion and management alternatives on net primary productivity in Inner Mongolia grassland, China. Global and Planetary Change, 2013, 108: 29-41.
[38] Chen S, Bai Y, Zhang L,etal. Comparing physiological responses of two dominant grass species to nitrogen addition in Xilin River Basin of China. Environmental and Experimental Botany, 2005, 53(1): 65-75.
[39] Chen S, Chen J, Lin G,etal. Energy balance and partition in Inner Mongolia steppe ecosystems with different land use types. Agricultural and Forest Meteorology, 2009, 149(11): 1800-1809.
[40] Borer E T, Seabloom E W, Gruner D S,etal. Herbivores and nutrients control grassland plant diversity via light limitation. Nature, 2014, 508(7497): 517-520.
[41] Shi Y, Wang Z Q, Zhang X Y,etal. Effects of nitrogen and phosphorus addition on soil microbial community composition in temperate typical grassland in Inner Mongolia. Acta Ecologica Sinica, 2014, 34(17): 4943-4949.
施瑶, 王忠强, 张心昱, 等. 氮磷添加对内蒙古温带典型草原土壤微生物群落结构的影响. 生态学报, 2014, 34(17): 4943-4949.
[42] Stevens M H, Carson W P. Plant density determines species richness along an experimental fertility gradient. Ecology, 1999, 80(2): 455-465.
[43] Hautier Y, Niklaus P A, Hector A. Competition for light causes plant biodiversity loss after eutrophication. Science, 2009, 324(5927): 636-638.
[44] Clark C M, Tilman D. Recovery of plant diversity following N cessation: effects of recruitment, litter, and elevated N cycling. Ecology, 2010, 91(12): 3620-3630.
