开食料中不同NDF水平对犊牛生长性能、瘤胃内环境及血清生化指标的影响
2018-05-22任春燕毕研亮杜汉昌于博屠焰郭艳丽刁其玉
任春燕,毕研亮,杜汉昌,于博,屠焰,郭艳丽,刁其玉*
(1.甘肃农业大学动物科学技术学院,甘肃 兰州 730070;2.中国农业科学院饲料研究所,农业部饲料生物技术重点试验室,北京 100081;3.奶牛营养学北京市重点实验室,北京 100081;4.山东银香伟业集团有限公司,山东 菏泽274400)
在反刍动物日粮与营养中,精饲料和粗饲料的配比为一个很有意义的参数,大致上给出了动物的日粮营养素水平。传统的犊牛饲喂方式通过提供定量的液态饲料如鲜牛奶和代乳品奶,鼓励采食固体饲料,以促进瘤胃发育和早期断奶。通常精饲料比粗饲料更利于刺激瘤胃发育[1]。在瘤胃发酵中提供精饲料可以发酵碳水化合物产生挥发性脂肪酸(VFA)进而刺激瘤胃乳头发育[1-2]。但是犊牛只饲喂高水平易发酵碳水化合物会降低瘤胃pH[3]和引起瘤胃乳头角质化,会严重损害胃肠功能,影响饲料利用效率以及犊牛的健康和福利[4]。而犊牛饲喂粗料可以促进瘤胃肌肉发育,维持上皮细胞完整性,促进瘤胃pH和瘤胃容积增大[5],防止瘤胃乳头凝集和分支[6],对瘤胃内环境产生积极影响,进而促进犊牛生产性能和健康状况[5,7]。已有研究表明,犊牛早期饲喂粗料利于犊牛瘤胃发酵和瘤胃发育[8],可以提高固体饲料采食量和生长性能[9-10]。
开食料中补充粗饲料对犊牛影响的研究结果不尽相同。一些研究人员认为在开食料中补充粗饲料可降低生产性能[11],可能是因为减少了饲粮能量密度[12]。然而其他研究人员认为在开食料中补充粗饲料可促进开食料采食量和饲料利用率[3,5]。Nemati等[13]认为粗饲料对犊牛的生长性能受粗饲料水平、来源、物理形态以及饲喂方式等因素的影响,其中粗饲料的添加水平被认为影响犊牛生长性能最重要的因素,而且认为补充粗饲料可以增加断奶后干物质采食量[9]。但是在饲喂一定来源的粗饲料情况下,犊牛开食料中最适的添加水平还未确定。因此,本研究拟通过评价粗饲料和精饲料混合制粒的开食料中不同中性洗涤纤维(neutral detergent fiber, NDF)水平对犊牛的开食料采食量、日增重、挥发性脂肪酸组成和血清生化指标的影响,为犊牛开食料中适宜NDF添加水平的确定提供数据支持,为犊牛开食料的合理配制,提高奶牛养殖业经济效益提供理论依据。
1 材料与方法
1.1 试验时间和地点
试验于2017年4-9月在山东银香伟业有限公司第二牧场开展。
1.2 试验设计
选用初生重为(42±2.5) kg,饲喂足量初乳的中国荷斯坦犊牛60头,其中公犊牛36头,母犊牛24头。采用完全随机区组设计,随机分为4组(A、B、C和D),每组15头,其中9头公犊牛,6头母犊牛,试验设计见表1。
1日龄开始,犊牛每日饲喂巴氏杀菌牛奶2次(6:30和17:00),1~28日龄饲喂5 L·d-1,28~65日龄饲喂8 L·d-1,65日龄后减至4 L·d-1,70日龄断奶。15日龄开始,4个处理组分别饲喂NDF水平为10%、15%、20%和25%的开食料,每日饲喂2次(7:00和16:30),保证料盆每日有剩料,自由饮水,试验期112 d。
1.3 试验日粮
分别饲喂4种含有不同NDF(NDF来源为苜蓿干草和燕麦干草,表2)水平的颗粒料。日粮制成颗粒(直径为6 mm)。通过调整各成分添加比例使蛋白质水平保持一致,开食料营养组成及营养水平见表2。
1.4 饲养管理
试验开始前对所有犊牛岛用消毒剂进行全面清洗消毒,晾晒之后使用。所有试验犊牛出生后进行正常免疫程序,于犊牛岛单独饲养。每日清晨饲喂后更换犊牛岛垫料,保证犊牛岛干燥清洁。整个试验期自由饮用清洁水。
1.5 测定指标及方法
1.5.1采食量和料重比的测定 试验第15天开始,详细记录每天每头犊牛的投料量和剩料量,计算开食料采食量。试验期间每2周晨饲前空腹测定每头犊牛的体重并计算每组日增重(average daily gain, ADG)和饲料转化率(feed conversion ratio, G∶F)。
1.5.2瘤胃液的采集和处理 正式试验第90天(断奶后20 d),每组选取6头犊牛,在晨饲后1~2 h通过口腔采液器从瘤胃采集50 mL瘤胃液样(舍弃最初采集的30 mL含有唾液的瘤胃液),立即用便携式pH计测定其pH,然后分装于10 mL的离心管中,于-20 ℃保存。测定瘤胃液氨态氮(NH3-N)、挥发性脂肪酸(VFA)浓度,计算总挥发性脂肪酸浓度(total volatile fatty acid,TVFA)及每种挥发酸的比例。
1.5.3血清生化指标 正式试验第90天,每组选取6头犊牛于晨饲前1 h颈静脉采血10 mL,3000 r·min-1离心20 min,收集血清分装于1.5 mL离心管中,-20 ℃下保存。用全自动生化分析仪测定血清总蛋白、葡萄糖和尿素氮。

表1 试验设计Table 1 The experiment design
1.6 统计分析
采用SAS 9.1统计软件中的方差分析(one-way ANOVA)和关于重复测量数据的MIXED模型进行分析。统计分析以P<0.05为差异显著,P<0.01为差异极显著,P在0.05~0.10为有提高或降低的趋势。
2 结果与分析
2.1 开食料中不同NDF水平对犊牛日增重和饲料转化率的影响
由表3可知,15~42日龄和70~112日龄B组日增重均高于A、C和D组,分别高出45.69、39.79和117.13 g·d-1(P<0.05),35.33、153.29和145.93 g·d-1(P<0.05)。70~112日龄,B、C和D组开食料采食量显著高于A组,分别高出255.20、252.48和392.27 g·d-1。A和B组犊牛的饲料转化率显著高于C和D组(P<0.05)。

表4 开食料中不同NDF水平对犊牛瘤胃液pH和NH3-N的影响Table 4 Effects of starter diets with different NDF levels on pH and NH3-N of rumen fluid in calves
2.2 开食料中不同NDF水平对犊牛瘤胃液pH和NH3-N含量的影响
由表4可知,瘤胃液pH在35和70日龄时C组显著高于其他3组(P<0.05);112日龄,A(6.22)和B(6.25)组显著高于C(5.55)和D组(5.83)(P<0.01)。NH3-N含量在90日龄前差异不显著;112日龄时,B、C和D组较A组显著降低16.72%、44.19%和52.10%(P<0.05)。
2.3 开食料中不同NDF水平对犊牛瘤胃液挥发性脂肪酸的影响
由表5可知,总挥发性脂肪酸含量随NDF水平的提高先升高后降低。C和D组乙酸比例显著高于A和B组(P<0.05)。B组丙酸比例较A、C和 D组提高4.24%、3.18%和6.43%(P<0.05),乙酸/丙酸显著低于D组(P<0.05)。丁酸和戊酸比例随NDF水平的提高有降低的趋势,但差异不显著(P>0.05)。
2.4 开食料中不同NDF水平对犊牛血清生化指标的影响
由表6可知,各处理组间血清TP、GLU和血清UN含量无显著差异(P>0.05)。B组血清TP含量略高于其他3个处理组,但差异不显著(P>0.05)。C组血清UN含量高于其他3个处理组,但差异不显著(P>0.05)。
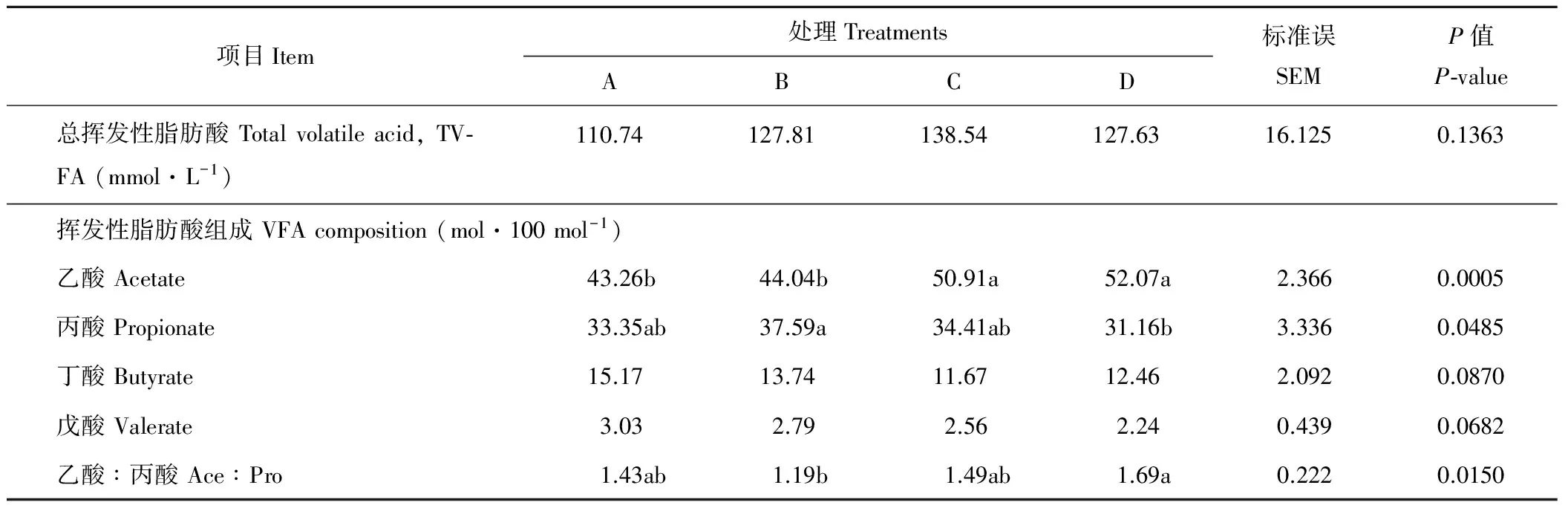
表5 开食料中不同NDF水平对犊牛90日龄(断奶后20 d)瘤胃液挥发性脂肪酸的影响Table 5 Effects of starter diets with different NDF on the VFA of rumen fluid 90 d of aged (20 d after weaning) in calves
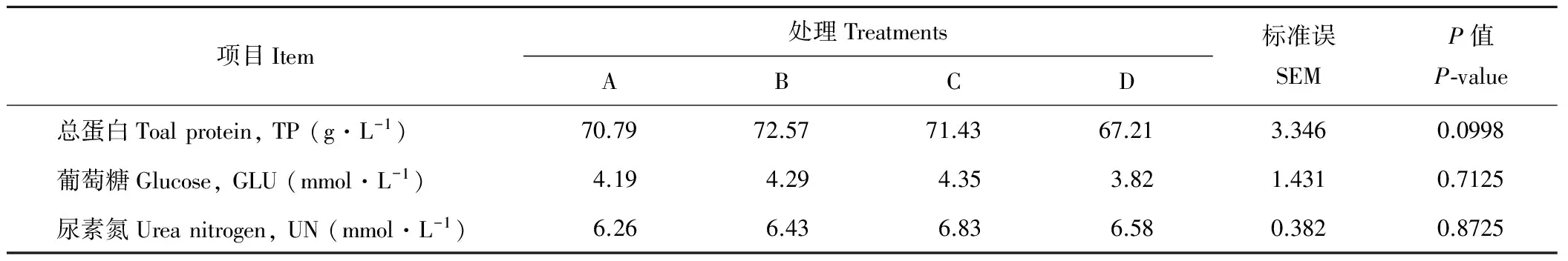
表6 开食料中不同NDF水平对犊牛血清生化指标的影响Table 6 Effects of starter diets with different NDF levels on serum biochemical parameters in calves
3 讨论
3.1 开食料中不同NDF水平对犊牛日增重和饲料转化率的影响
本试验中随犊牛日龄的增加,开食料采食量显著增加,ADG随着增加。42~70日龄各处理组间犊牛ADG无显著差异,可能是因为此阶段犊牛以牛奶为主要能量来源,开食料采食量较低,所以对ADG影响较小。Nemati等[13]认为犊牛ADG受苜蓿干草添加水平的影响,当补充苜蓿达到25%时(24.5%NDF)断奶后ADG增加。另外,通过向饲粮中添加切碎的秸秆[14]或棉籽粕[15],增加了饲粮NDF的水平,提高了犊牛断奶前后开食料采食量和体增重。本试验中15%NDF组断奶后ADG明显增加,与Kosiorowska等[16]的研究结果相一致,因为适宜的NDF水平和淀粉含量,利于改善瘤胃内环境,进而促进了犊牛ADG的增加[12]。另有研究人员认为补充粗料使体重和ADG增加,是由于粗饲料使胃肠容积填充和胃肠组织的重量增加[17-18],但是在本试验中发现,15%组瘤胃重显著高于其他3个处理组,而且20%和25%组并没有因为NDF水平增加而体重增加。因此,可以推导出,胃肠容积填充并不是影响体重和ADG的主要因素,而是胃肠组织的发育对体重和ADG的变化起主要作用。断奶后(70~112 d)各处理组间开食料采食量差异显著,其中15%组开食料采食量高于其他3组,可能是由于适宜的NDF水平促进了瘤网胃的容积和发育[10,16],进而增加了开食料采食量。 EbnAli等[7]以不同方式向犊牛添加苜蓿(15.8%~17.3%NDF)颗粒,促进了断奶后犊牛的开食料采食量。试验全期10%和15%NDF组饲粮转化率得到显著改善,较20%和25%NDF组提高了10.96%,提高了饲料利用效率。因此,向1~3月龄犊牛提供以苜蓿和燕麦干草为来源的15%NDF有利于提高犊牛生产性能。
3.2 开食料中不同NDF水平对犊牛瘤胃液pH和NH3-N的影响
瘤胃pH对反刍动物瘤胃发育和瘤胃发酵,以及整个生命健康都起着至关重要的作用。一般瘤胃液pH受VFA发酵和吸收速率的影响,而后者又受瘤胃内容物消化速率和缓冲能力的影响[18]。本试验中未出现补饲干草与瘤胃pH呈正相关的结果与Laarman等[19]报道结果不一致,可能是由于不同的粗饲料来源及不同NDF水平对犊牛反刍和唾液分泌的刺激作用不同引起的[20]。Terré等[17]发现颗粒料NDF水平对断奶后瘤胃pH值无影响,本试验90日龄瘤胃pH与此相一致。瘤胃pH过低会影响瘤胃微生物的组成,使部分微生物发生迁移,降低饲料的消化利用[20]。112日龄,20%(5.55)和25%(5.83)组瘤胃液pH显著低于10%(6.22)和15%(6.25)组,可能是断奶后犊牛采食量增加,NDF摄入量相应增加,而瘤胃上皮尚未发育完全,使挥发酸的产量超过了瘤胃上皮细胞的吸收能力,引起瘤胃液pH降低[18]。
NH3-N是瘤胃主要的代谢物,是瘤胃功能发育的标志。瘤胃中NH3-N既是饲料蛋白质、内源性蛋白质和非蛋白氮分解的终产物,同时也是瘤胃微生物合成微生物蛋白质(microprotein,MCP)的主要氮源,一定程度上可以反映出瘤胃微生物分解含氮物质产生NH3及对其摄取利用的情况[21]。Agle等[22]研究表明,奶牛采食高粗料日粮时会引起瘤胃中NH3-N水平的降低,本研究结果与此相一致。杨宏波等[23]认为瘤胃微生物的生长,其NH3-N浓度不宜过高或过低,保持最适浓度的NH3-N是保证瘤胃微生物蛋白产量最重要的条件。Hristov等[24]认为NH3-N浓度过低会限制MCP的合成,浓度过高会使瘤胃微生物降解氮源释放氨气(NH3)的速率超过微生物利用NH3合成MCP的速率,进而抑制微生物对NH3-N的利用,造成瘤胃氮素循环中N的损失。本试验中,10%NDF组NH3-N浓度显著高于20%和25%NDF组,因此在犊牛开食料中补充15%NDF会具有适宜的NH3-N浓度,利于提高瘤胃代谢活性,促进动物生长性能。
3.3 开食料中不同NDF水平对犊牛瘤胃液挥发性脂肪酸比例的影响
瘤胃挥发性脂肪酸(VFA)为反刍动物能量利用中一个重要的中间代谢产物,其含量及组成比例是反映瘤胃消化代谢活动的重要指标。本试验TVFA浓度为67~160 mmol·L-1,在Castells等[25]的30~160 mmol·L-1范围。Baldwin等[26]研究表明,瘤胃发酵和TVFA的产量与固体饲料和NDF的采食量有关。Terré等[17]发现断奶后高NDF(26.7%NDF)颗粒料组TVFA含量高于低NDF组(18.2%NDF)。本试验中,断奶后随开食料NDF水平的提高,瘤胃TVFA含量先升高后降低,与采食量的变化基本一致。可能因为断奶后瘤胃微生物区系的建立逐步完善,开食料采食量增加,粗饲料发酵利用率高,促进了TVFA含量的升高。
犊牛瘤胃发酵参数受粗饲料的来源、水平及饲喂方式的影响。Thomas等[15]用一种未添加棉籽粕(25.8%NDF),另一种添加棉籽粕(38.6%NDF)的开食料饲喂犊牛发现对瘤胃挥发酸比例无显著影响。Quigley等[27]研究发现,与未补充苜蓿干草的犊牛相比,补充苜蓿干草犊牛丙酸比例增加且丁酸比例降低。Terré等[17]发现,与未补充粗饲料的犊牛相比,饲喂粗饲料犊牛乙酸比例较高,丁酸和戊酸比例较低,本研究结果与此相一致。其中丙酸和丁酸被认为是刺激瘤胃上皮发育的最主要挥发酸[26]。本研究中,15%NDF组丙酸比例高于其他3个处理组。很多文献显示饲粮淀粉含量高时,瘤胃分解淀粉的微生物占优势,丙酸浓度显著增加。Sutton等[28]研究证明精料中增加淀粉的含量也可使瘤胃液中丙酸浓度增加,因此也可能是由于15%NDF组具有适宜的淀粉比例,增加丙酸浓度。而丙酸又是反刍动物体内糖异生的主要前提物质,因此促进丙酸的生成有利于提高犊牛生产性能。Cline等[29]研究发现,微生物生长和纤维降解与戊酸比例之间呈正相关关系。本试验随NDF水平升高,戊酸比例降低,说明适宜的NDF水平利于纤维分解菌生长,这与Kosiorowska等[16]向犊牛补充适宜NDF的开食料,可以改善纤维素分解菌的生长相一致。也可能由于犊牛饲喂高NDF饲粮瘤胃A/P提高,说明瘤胃乙酸比例增加,相反补充粗饲料A/P降低主要是由于瘤胃丙酸比例增加[30]。本试验中15%NDF组的A/P在90日龄时显著低于其他处理组,由此说明15%NDF组属于丙酸发酵。
3.4 开食料中不同NDF水平对犊牛血清生化指标的影响
血清中各种生化成分是动物体生命活动的物质基础,其含量及变化规律是动物体重要的生物学特征[31]。血清TP是机体蛋白质合成代谢的一个重要指标,其含量高低可以反映蛋白质的代谢情况,当血清TP含量升高,表明动物的新陈代谢加强,利于动物生长[32-33]。本试验中在饲粮蛋白质水平相近的情况下,15%NDF组血清TP含量略高于其他3个处理组,说明开食料中补充适宜的NDF利于促进犊牛生长发育。
血清GLU含量对于犊牛各组织器官的生理功能是极其重要的,是各组织细胞活动的主要能量来源[34]。断奶后犊牛不能或很少直接消化饲粮中碳水化合物获得机体所需GLU,而是通过糖异生途径主要由丙酸合成[35]。一般GLU含量低于6.1 mmol·L-1[36],本试验中GLU含量在3.82~4.35 mmol·L-1,组间差异不显著。血清UN是蛋白质分解的最终产物,可以较准确地反映体内蛋白质的代谢状况和饲粮氨基酸的平衡情况,蛋白质代谢良好时,血清UN含量较低,说明机体对饲料蛋白质的利用率提高[37]。本试验中4个处理组间血清UN含量无显著差异,但15%NDF组略低于10%、20%和25%NDF组,说明开食料中添加15%NDF有利于提高犊牛对开食料蛋白质的利用率。
4 结论
1)日粮中的NDF水平对1~3月龄犊牛断奶后日增重、饲料转化率和瘤胃内环境均产生影响,对犊牛血清生化指标无影响。
2)提供以苜蓿和燕麦草为主要NDF来源的开食料中的最适NDF水平为15%左右。
参考文献References:
[1] Lesmeister K E, Heinrichs A J. Effects of corn processing on growth characteristics, rumen development and rumen parameters in neo-natal dairy calves. Journal of Dairy Science, 2004, 87(10): 3439-3450.
[2] Khan M A, Lee H J, Lee W S,etal. Structural growth, rumen development, metabolic and immune response of Holstein male calves fed milk through step-down and conventional methods. Journal of Dairy Science, 2007, 90(7): 3376-3387.
[3] Daneshvar D M, Khorvash E, Ghasemi,etal. The effect of restricted milk feeding through conventional or step-down methods with or without forage provision in starter feed on performance of Holstein bull calves. Journal of Animal Science, 2015, 93(8): 3979-3989.
[4] Bull L S, Bush L J, Friend J D,etal. Incidence of ruminal parakeratosis in calves fed different rations and its relation to volatile fatty acid absorption. Journal of Dairy Science, 1965, 48(11): 1459-1466.
[5] Beiranvand H, Ghorbani G R, Khorvash M,etal. Interactions of alfalfa hay and sodium propionate on dairy calf performance and rumen development. Journal of Dairy Science, 2014, 97(4): 2270-2280.
[6] Greenwood R H, Morril J L, Titgemeyer E C,etal. A new method of measuring diet abrasion and its effect on the development of the forestomach. Journal of Dairy Science, 1997, 80(10): 2534-2541.
[7] EbnAli A, Khorvash M, Ghorbani G R,etal. Effects of forage offering method on performance, rumen fermentation, nutrient digestibility, blood metabolites, and nutritional behavior in Holstein dairy calves. Journal of Animal Physiology and Animal Nutrition, 2016, 100(5): 820-827.
[8] Mirzaei M, Khorvash M, Ghorbani G R,etal. Effects of supplementation level and particle size of alfalfa hay on growth characteristics and rumen development in dairy calves. Journal of Animal Physiology, 2015, 99(3): 553-564.
[9] Castells L, Bach A, Araujo G,etal. Effect of different forage sources on performance and feeding behavior of Holstein calves. Journal of Dairy Science, 2012, 95(1): 286-293.
[10] Montoro C E, Miller-Cushon K, DeVries T J,etal. Effect of physical form of forage on performance, feeding behavior, and digestibility of Holstein calves. Journal of Dairy Science, 2013, 96(2): 1117-1124.
[11] Hill T M, Bateman H G, Aldrich J M,etal. Roughage amount, source, and processing for diets fed to weaned dairy calve. Journal of Animal Science, 2010, 26(2): 181-187.
[12] Drackley J K. Calf nutrition from birth to breeding. Veterinary Clinics of North America: Food Animal Practice, 2008, 24(1): 55-86.
[13] Nemati M, Amanlou H, Khorvash M,etal. Effect of different alfalfa hay levels on growth performance, rumen fermentation, and structural growth of Holstein dairy calves. Journal of Animal Science, 2016, 94(3): 1141-1148.
[14] Thomas D B, Hinks C E. The effect of changing the physical form of roughage on the performance of the early-weaned calf. Animal Production, 2015, 35(3): 375-384.
[15] Thomas D B, Hinks C E. The effect of changing the physical form of roughage on the performance of the early-weaned calf. Animal Production, 2015, 35(3): 375-384.
[16] Kosiorowska A, PuggaardL M, Hedemann S,etal. Gastrointestinal development of dairy calves fed low- or high-starch concentrate at two milk allowances. Animal, 2011, 5(2): 211-219.
[17] Terré M, Pedrals E, Dalmau A,etal. What do preweaned and weaned calves need in the diet: A high fiber content or a forage source. Journal of Dairy Science, 2013, 96(8): 5217-5225.
[18] Williams P E V, Fallon R J, Innes G M,etal. Effect on food intake, rumen development and live weight of calves of replacing barley with sugar beet-citrus pulp in a starter diet. Animal Production Science, 1987, 44(1): 65-73.
[19] Laarman A H, Oba M. Short communication: effect of calf starter on rumen pH of Holstein dairy calves at weaning. Journal of Dairy Science, 2011, 94(11): 5661-5666.
[20] Penner G B, Oba M. Increasing dietary sugar concentration may improve dry matter intake, ruminal fermentation, and productivity of dairy cows in the postpartum phase of the transition period. Journal of Dairy Science, 2009, 92(7): 3341-3353.
[21] Liu J, Diao Q Y, Zhao Y G,etal. Effects of dietary NFC/NDF ratios on rumen pH, NH3-N and VFA of meat sheep. Chinese Journal of Animal Nutrition, 2012, 24(6): 1069-1077.
刘洁, 刁其玉, 赵一广, 等. 饲粮不同NFC/NDF对肉用绵羊瘤胃pH、氨态氮和挥发性脂肪酸的影响. 动物营养学报, 2012, 24(6): 1069-1077.
[22] Agle M, Hristov A, Zaman S,etal. Effect of dietary concentrate on rumen fermentation, digestibility and nitrogen losses in dairy cows. Journal of Dairy Science, 2010, 93(9): 4211-4222.
[23] Yang H B, Liu H, Zhan J S,etal. Effects of diet pellets with different concentrate-roughage ratios on rumen fermentation parameters and microorganism abundance in weaned bull calves. Acta Prataculturae Sinica, 2015, 24(12): 131-138.
杨宏波, 刘红, 占今舜, 等. 不同精粗比颗粒饲料对断奶公犊牛瘤胃发酵参数和微生物的影响. 草业学报, 2015, 24(12): 131-138.
[24] Hristov A N, Broderick G A. In vitro determination of ruminal protein degradability using [15N] ammonia to correct for microbial nitrogen uptake. Journal of Animal Science, 1994, 72(5): 1344-1354.
[25] Castells L, Bach A, Aris A,etal. Effects of forage provision to young calves on rumen fermentation and development of the gastroin-testinal tract. Journal Dairy Science, 2013, 96(8): 5226-5236.
[26] Baldwin VI R L, McLeod K R, Klotz J L,etal. Rumen development, intestinal growth and hepatic metabolism in the pre- and post-weaning ruminant. Journal of Dairy Science, 2004, 87(1): 55-65.
[27] Quigley J D, Steen T M, Boehms S I. Postprandial changes of selected blood and ruminal metabolites in ruminating calves fed diets with or without hay. Journal of Dairy Science, 1992, 75(1): 228-235.
[28] Sutton J D, Morant S V, Bines J A,etal. Effect of altering the starch: fiber ratio in the concentration on hay intake and milk production by Friesian cows. Journal of Agricultural Science, 1993, 120(3): 379-390.
[29] Cline J H, Hershberger T V, Bentley O G,etal. Utilization and/or synthesis of valeric acid during the digestion of glucose, starch and cellulose by rumen micro-organisms in vitro. Journal Animal Science, 1958, 17(2): 284-292.
[30] Khan M A, Weary D M, von Keyerslingk M A G. Hay intake improves performance and rumen development of calves fed higher quantities of milk. Journal of Dairy Science, 2011, 94(7): 3547-3553.
[31] Wang Z G. The influence of different dietary riboflavin concentration on the immune performance and blood biochemical parameters of New Yangzhou Chickens. Yangzhou: Yangzhou University, 2005.
汪张贵. 日粮核黄素添加水平对新扬州仔鸡免疫机能和血液生化指标影响的研究. 扬州: 扬州大学, 2005.
[32] Chen W, Guo Y M, Huang Y Q. Effects of supplementation of corn oil and lard on growth and slaughter performance, and serum bio-chemical indices in broilers. Chinese Journal of Animal Nutrition, 2011, 23(8): 1101-1108.
陈文, 呙于明, 黄艳群. 玉米油和猪油对肉鸡生产性能、屠宰性能及血清生化指标的影响. 动物营养学报, 2011, 23(8): 1101-1108.
[33] Wang Y H, Wang C Z, Gao Y G,etal. Effects of alfalfa saponins on the nutrient digestibility, production performance and blood bio-chemical index of weaned piglets. Acta Prataculturae Sinica, 2009, 18(5): 115-122.
王彦华, 王成章, 高永革, 等. 苜蓿皂苷对断奶仔猪生产性能、消化率及血液生化指标的影响. 草业学报, 2009, 18(5): 115-122.
[34] Zhou Y, Diao Q Y, Tu Y,etal. Effects of yeast beta-glucan on the production performance and physiological and biochemical blood indexes of early weaning calves. Chinese Journal of Animal Science, 2010, 46(13): 48-51.
周怿, 刁其玉, 屠焰, 等. 酵母β-葡聚糖对早期断奶犊牛生产性能和血液生理生化指标的影响. 中国畜牧杂志, 2010, 46(13): 48-51.
[35] Cui X, Diao Q Y, Zhang N F,etal. Effects of diets with different energy levels on growth and development and serum indexes of salves aged from 3 to 6 months. Chinese Journal of Animal Nutrition, 2014, 26(4): 947-961.
崔祥, 刁其玉, 张乃锋, 等. 不同水平的饲粮对 3~6 月龄犊牛生长发育及血清指标的影响. 动物营养学报, 2014, 26(4): 947-961.
[36] Wang D, Li F D, Zhang Y D,etal. Mixed silage of potato pulp and corn straw affect growth performance, rumen environments and blood biochemical parameters of mutton sheep. Acta Prataculturae Sinica, 2012, 21(5): 47-54.
王典, 李发弟, 张养东, 等. 马铃薯淀粉渣-玉米秸秆混合青贮料对肉羊生产性能、瘤胃内环境和血液生化指标的影响. 草业学报, 2012, 21(5): 47-54.
[37] Stanley C C, Williams C C, Jenny B F,etal. Effects of feeding milk replacer once versus twice daily on glucose metabolism in Holstein and Jersey calves. Journal of Dairy Science, 2002, 85(9): 2335-2343.
王迎新, 陈先江, 娄珊宁, 等. 草原灌丛化入侵:过程、机制和效应. 草业学报, 2018, 27(5): 219-227.
Wang Y X, Chen X J, Lou S N,etal. Woody-plant encroachment in grasslands: a review of mechanisms and aftereffects. Acta Prataculturae Sinica, 2018, 27(5): 219-227.
