两个豚鼠品系眼球生物学特性比较及视网膜近视相关机制研究
2018-05-04陈雁虹卫振艾志鹏刘迪文
陈雁虹,卫振,艾志鹏,刘迪文
(浙江大学实验动物中心,杭州 310058)
近视是指眼在不使用调节时,平行光线通过眼的屈光系统屈折后,焦点落在视网膜之前的一种屈光状态。在亚洲进行的近视流行病学调查结果显示中国人近视率普遍高于白色人种[1],尤其是中国大陆、中国台湾、中国香港和新加坡青少年的近视发病率高于世界其他地区。针对近视患病率急剧升高的现状,科学家们已陆续开展对近视病因和发病机制的研究。20世纪70年代近视动物模型的建立,为近视研究开辟了一条新的途径,一些无法在人类身上进行的研究可以通过动物模型来实现。合适的近视动物模型将直接决定实验研究是否成功和实验结果是否准确。
豚鼠因其出生后即开眼,眼球的发育过程与人类相似,视力优于大小鼠,温顺配合好,易获得测量数据等优势,而成为目前最常用于近视疾病模型研究的实验动物。Zmu-1:DHP豚鼠是浙江大学实验动物中心经过近二十年悉心培育获得的近交系品系[2],在该品系豚鼠用于眼科实验的过程中发现,该纯白色豚鼠品系与DHP豚鼠品系相比,具有易自发性近视的特点[3-4]。因此,作者选择将Zmu-1:DHP和DHP两个豚鼠品系进行眼球相关生物学特性比较,并对视网膜近视相关机制展开研究。
视黄酸(retinoic acid,RA)能够在视网膜细胞内合成,是影响巩膜发育、光感受器细胞分化、成熟的重要信使分子[5],视黄醛脱氢酶(retinal dehydrogenase,RALDH)和醛脱氢酶(aldehyde dehydrogenase,ALDH)是RA合成酶[6]。而酪氨酸羟化酶(tyrosine hydroxylase,TH)、酪氨酸激酶(tyrosine kinases,TK)、诱导型NO合成酶(inducible nitric oxide synthase,iNOS)、神经型NO合成酶(neuronal nitric oxide synthase,nNOS)、碱性成纤维细胞生长因子(basic fibroblast cell growth factor,bFGF)和转化生长因子β(transforming growth factor β,TGFβ)是已知参与调控近视发生发展的视网膜信号因子[7],通过对这些因子mRNA的表达检测,研究Zmu-1:DHP易自发性近视的机制。
1 材料与方法
1.1 材料
1.1.1 实验动物
3周龄清洁级Zmu-1:DHP豚鼠327只和DHP豚鼠100只,体重140~180 g,由浙江大学实验动物中心提供【SCXK(浙)2012-0052】,动物饲养在浙江大学实验动物中心清洁级屏障【SYXK(浙)2012-0178】。动物实验方案经浙江大学实验动物福利伦理审查委员会审查通过【ZJU2015-496-04】。
1.1.2 试剂与仪器
动物组织RNA提取和实时荧光定量试剂盒购自TaKaRa公司;引物由赛默飞世尔科技(中国)有限公司合成;眼球固定液由浙江大学医学院公共技术平台提供。
红外偏心摄影验光仪(eccentric infrared photoretinoscope,EIR),实验室自行搭建;角膜曲率计(OM.4,Topcon,Japan),实验室改良;A超测定仪(AVISO Echograph Class I-Type Bat,Quantel Medical,Clermont-Ferrand,France);脱水机(ASP300S,Leica,Germany),切片机(RM2235,Leica,Germany),摊烤片机(TEC-2500,Hestion),包埋机(EC350-1,Microm,Germany);倒置显微镜(U-25ND6,Olympus,Japan);CFX Connect实时荧光定量PCR仪(Bio-Rad,USA)。
1.2 方法
1.2.1 屈光度、角膜曲率与眼轴长度的测定[8]
豚鼠置于暗室检测台上,将待测眼对准验光仪的光源,调整豚鼠位置使Purkinje影像位于眼球瞳孔中央,待瞳孔和屈光状态稳定时,记录屈光值。每次连续测量三次,取平均值。
由助手固定豚鼠,操作改良角膜曲率计,使光标清晰并呈正圆形,然后调整曲率计进行读数。每只眼连续测量3次,记录水平子午线的角膜曲率度数,测量时间控制在5 min内。豚鼠眼睛滴入一滴0.5%盐酸丙美卡因滴眼液(Alcon,Puurs,Belgium),使用A超探头测量眼轴长度。
327只Zmu-1:DHP豚鼠和100只DHP豚鼠3周龄时通过屈光度测定,统计自发性近视发生率,并从筛选出的295只自发性近视Zmu-1:DHP豚鼠与100只DHP豚鼠中随机选取各12只豚鼠分别在4、6、8、10、12周时进行屈光度、角膜曲率与眼轴长度的连续测定。
1.2.2 视网膜组织结构的观察
12周时,处死Zmu-1:DHP豚鼠和DHP豚鼠各6只,取出一侧眼球,固定于眼球固定液中,进行常规病理取材、制片和HE染色,显微镜下进行组织结构观察。
1.2.3 实时PCR检测视网膜信号因子mRNA的表达
12周时,处死Zmu-1:DHP豚鼠和DHP豚鼠各6只,豚鼠视网膜剥离后,按照说明书提取视网膜总RNA,逆转录获得cDNA,在实时PCR专用96孔板中配制标准反应体系,进行SYBR Green I实时荧光定量PCR扩增,95℃孵育3 min,95℃ 10 s,60℃ 32 s,40个循环,融解曲线分析:95℃ 10 s,65℃ 5 s。各引物序列见表1。用基因相对定量法(2-ΔΔCt)计算目的基因相对表达。

表1 视网膜近视因子的PCR引物Tab.1 PCR primers of the myopia factors in retina
1.3 统计学分析
2 结果
2.1 两个豚鼠品系3周龄屈光度比较
3周龄时,327例Zmu-1:DHP豚鼠左、右眼屈光度平均值分别为-4.24和-6.27,以屈光度小于-0.25 D作为近视的标准,近视率达到90.21%,较DHP豚鼠左右眼屈光度平均值3.84和1.78,差异有显著性(P<0.01),见表2。
2.2 两个豚鼠品系4~12周屈光度、角膜曲率与眼轴长度变化比较
4~12周,Zmu-1:DHP豚鼠随周龄增长,左右眼近视度数均不断加深,而DHP豚鼠双眼均未近视,两品系间屈光度比较有显著性差异(P<0.01)。两个豚鼠品系左右眼的角膜曲率均随周龄增长而逐渐增大,但Zmu-1:DHP豚鼠较DHP豚鼠增长幅度较小,前者左眼第10、12周,右眼第12周角膜曲率明显小于后者(P<0.01)。4~12周,Zmu-1:DHP豚鼠眼轴长度从8.4 mm增长至约9.3 mm,DHP豚鼠眼轴长度从8.1 mm增长至约8.8 mm,前者眼轴长度大于后者,差异有显著性(P<0.01)。见表3。

表2 Zmu-1:DHP和DHP豚鼠3周龄屈光度比较Tab.2 Comparison of refraction between the Zmu-1:DHP and DHP guinea pigs at the age of three
注:L表示左眼,R表示右眼,**表示与DHP比较差异有显著性(P<0.01)。
Note. Symbol L means the left eye, R means the right eye;**P<0.01, compared with DHP guinea pigs.

表3 Zmu-1:DHP和DHP豚鼠4~12周屈光度、角膜曲率、眼轴长度变化比较Tab.3 Changes of refraction, corneal curvature and axial length at 4-12 weeks of age in the Zmu-1:DHP and DHP guinea =12)
注:L表示左眼,R表示右眼,**表示与DHP比较差异有显著性(P<0.01)。
Note. Symbol L means the left eye, R means the right eye;**P<0.01, compared with the DHP guinea pigs.
2.3 两个豚鼠品系视网膜组织结构比较
显微镜观察结果见图1。视网膜分为视神经纤维层、神经节细胞层、内丛状层、内核层、外丛状层、外核层、视杆与视锥层、色素上皮层。由图可见,两品系的视网膜分层清晰可见,Zmu-1:DHP豚鼠视网膜内核层较厚,外核层较薄,外核层细胞体积小,分布数目少;DHP豚鼠视网膜内核层较薄,而外核层较厚,外核层细胞体积大,分布数目较多(红色箭头示)。Zmu-1:DHP豚鼠色素上皮细胞层未见明显色素颗粒,而DHP豚鼠色素上皮细胞层内分布有大量棕黄色色素颗粒(蓝色箭头示)。同时Zmu-1:DHP豚鼠脉络膜较DHP豚鼠萎缩变薄(黄色箭头示)。
2.4 两个豚鼠品系视网膜信号因子mRNA表达差异
与DHP豚鼠比较,Zmu-1:DHP豚鼠视网膜TH的表达明显降低(P<0.01),TK的表达显著增强(P<0.01),iNOS、nNOS、bFGF和TGFβ的表达明显增强(P<0.05),而RALDH和ALDH虽然表达增强,但差异无显著性。见图2。
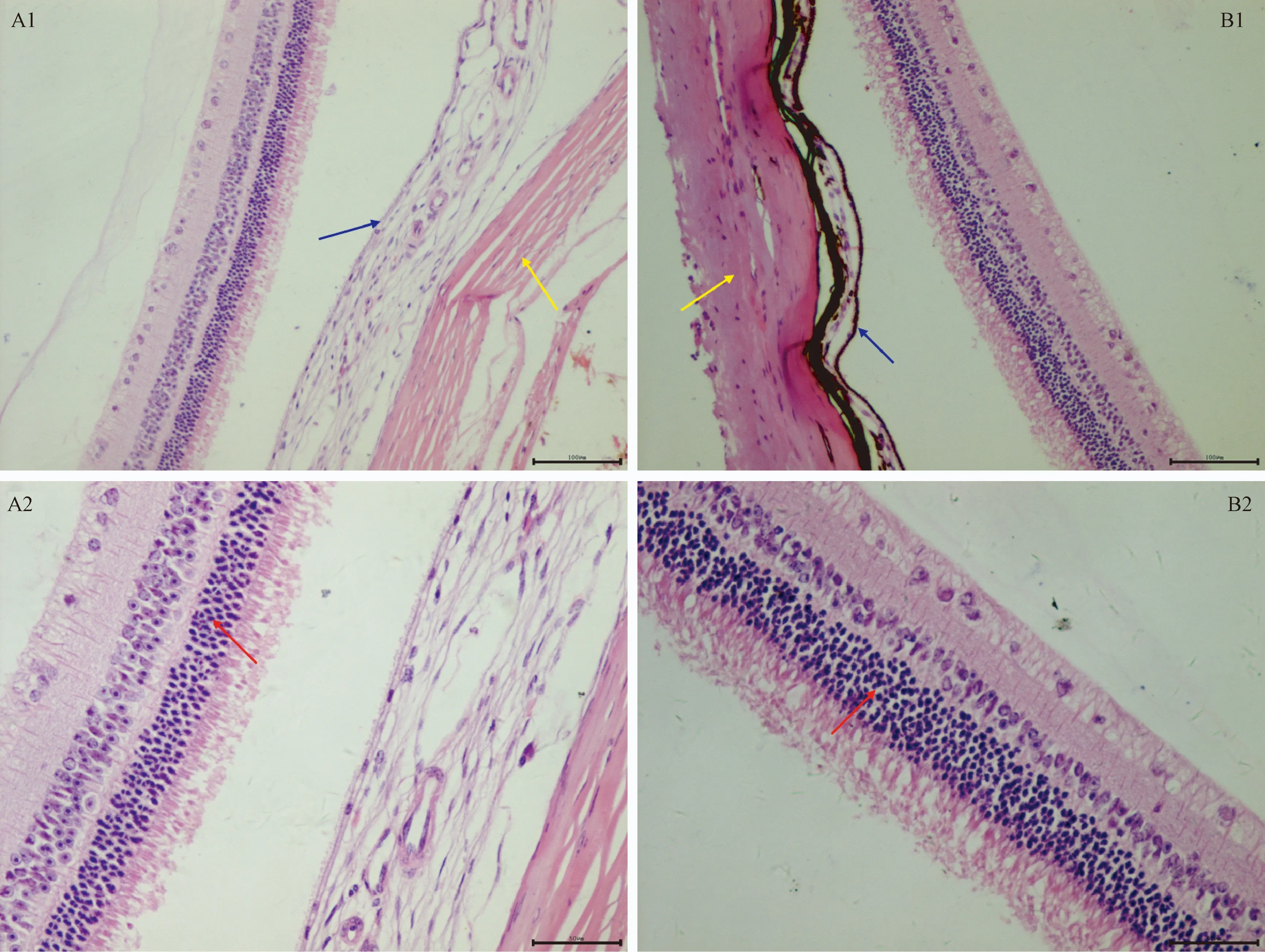
注:A: Zmu-1:DHP豚鼠;B: DHP豚鼠(1. HE染色,×100; 2. HE染色,×200)。红色箭头示视网膜外核层,蓝色箭头示色素上皮细胞层,黄色箭头示脉络膜。图1 Zmu-1:DHP和DHP豚鼠视网膜组织结构比较Note. A: Zmu-1:DHP guinea pigs; B: DHP guinea pigs (1. HE staining, ×100; 2. HE staining, ×200). The red arrow refers to retinal outer nuclear layer, the blue arrow refers to pigment epithelium, the yellow arrow refers to choroidal.Fig.1 Histologicl structure of the retina of the Zmu-1:DHP and DHP guinea pigs

注:*表示与DHP比较差异有显著性(P<0.05),**表示与DHP比较差异有显著性(P<0.01)。图2 Zmu-1:DHP和DHP豚鼠视网膜信号因子mRNA的表达差异Note.* P<0.05,** P<0.01, compared with the DHP guinea pigs.Fig.2 Myopia factors mRNA expression in the retina of Zmu-1:DHP and DHP guinea pigs
3 讨论
通常,在正常视觉环境下,大多数幼年动物的双眼均朝正视化方向发展,如普通DHP豚鼠,出生时呈远视状态, 随年龄增长,屈光状态逐渐调整,最终完成正视化过程。但达到正视化并非屈光度数为零,而是仍有一定的远视,这可能是豚鼠适应环境的一种表现。Zmu-1:DHP豚鼠却正好相反,出生时大部分呈近视状态,近视自发率达到90.21%,之后随年龄增加,近视不断加深,不能完成正视化过程。根据对Zmu-1:DHP豚鼠眼轴长度测量结果的分析,可以推测,该品系的近视是由于眼球前后径生长过快引起的轴性近视。光镜下观察视网膜组织切片,Zmu-1:DHP豚鼠脉络膜萎缩变,使得玻璃体腔深度增加,眼轴变长,视网膜成像焦点落后,从而导致近视,为上述推测提供了依据。人类近视的主要致病原因是遗传和环境[9],其中一种轴性近视即由于眼轴增长,脉络膜变薄而形成。Zmu-1:DHP豚鼠的自发性近视与人类的轴性近视极为相似,可以作为实验动物模型应用于人类近视机制的研究。
在近视的发生发展中,已知受到多种近视因子影响,而视网膜作为感知视觉的组织在其中起到了重要的作用。目前较为公认的机制是在视网膜调控下,巩膜基质金属蛋白酶2(matrix metalloproteinase 2,MMP-2)与其抑制因子组织金属蛋白酶抑制剂2(tissue inhibitor of matrix metalloproteinase 2,TIMP-2)二者表达失衡,巩膜主动重塑,引起眼轴过度延长。
本文通过对两个豚鼠品系视网膜信号因子RALDH、ALDH、TH、TK、iNOS、nNOS、bFGF、TGFβ的mRNA表达检测,发现,Zmu-1:DHP豚鼠RALDH、ALDH表达增强,可能导致视网膜合成RA增加,从而引起MMP-2/TIMP-2表达失衡,促使巩膜细胞外基质重塑;同时TH表达降低,TH是多巴胺(dopamine,DA)的合成酶,DA是黑色素合成的前体,其表达降低使得DA合成减少,含量下降,视网膜黑色素缺乏,导致进入眼内的光线增加,散射增多,视网膜成像质量下降,从而进一步引起视网膜合成、分泌其它信号因子,如TK、iNOS、nNOS、bFGF、TGFβ,这些信号因子共同作用,使得Zmu-1:DHP豚鼠形成自发性近视。
综上所述,Zmu-1:DHP豚鼠可以为眼科学科研工作者研究近视相关病理、生理变化过程及机制提供一个遗传背景清楚,近视发病率高,近视状态稳定的疾病动物模型选择,其自发性近视更多机制有待后续更进一步的研究。
参考文献(References)
[1] Seet B,Wong TY,Tan DT, et al. Myopia in Singapore:taking a public health approach [J]. Brit J Ophthalmol,2001, 85(5): 521-526.
[2] 刘迪文,谢敏,陈雁虹,等. Zmu-1:DHP近交系豚鼠的培育及其分子遗传结构初步鉴定 [J]. 中国实验动物学报,2017,25(1): 90-96.
Liu DW, Xie M, Chen YH, et al. Breeding of Zmu-1:DHP inbred strain guinea pig and preliminary analysis of molecular genetic structure of the strain [J]. Acta Lab Anim Sci Sin,2017,25(1): 90-96.
[3] Jiang L, Schaeffel F, Zhou X, et al. Spontaneous axial myopia and emmetropization in a strain of wild-type guinea pig(Cavia porcellus)[J]. Invest Ophthalmol Vis Sci. 2009,50(3): 1013-1019.
[4] Jiang L, Long K, Schaeffel F, et al. Disruption of emmetropization and high susceptibility to deprivation myopia in albino guinea pigs [J]. Invest Ophthalmol Vis Sci. 2011, 52(9): 6124-6132.
[5] Wang S, Liu S, Mao J, et al. Effect of retinoic acid on the tight junctions of the retinal pigment epithelium-choroid complex of guinea pigs with lens-induced myopia in vivo [J]. Int J Mol Med.2014, 33(4): 825-832.
[6] Rada JA, Hollaway LR, Lam W, et al. Identification of RALDH2 as a visually regulated retinoicacid synthesizing enzyme in the chick choroid [J]. Invest Ophthalmol Vis Sci. 2012, 53(3): 1649-1662.
[7] 毛俊峰, 刘双珍, 秦文娟, 等. PKC对豚鼠近视眼视网膜Müller细胞近视因子基因的调控作用 [J]. 眼科研究, 2009, 27(2): 86-90.
Mao JF, Liu SZ, Qin WJ, et al. The modulation effects of PKC on gene expression of myopic factors in retinal Müller cells in myopic eye of guinea pig [J]. Chin J Exp Ophthalmol, 2009, 27(2): 86-90.
[8] 卫振, 张森, 蒋丽琴, 等. 两品系豚鼠屈光状态和眼球径的发展和比较 [J]. 实验动物学报, 2016, 24(1): 92-96.
Wei Z, Zhang S, Jiang LQ, et al. Comparison of the refractive development and ocular growth between two strains of guinea pigs [J]. Acta Lab Anim Sci Sin,2016, 24(1): 92-96.
[9] Sun J, Zhou J, Zhao P, et al. High prevalence of myopia and high myopia in 5060 Chinese university students in Shanghai [J]. Invest Ophthalmol Vis Sci. 2012, 53(12): 7504-7509.
