模式生物海洋青鳉鱼的视觉结构与功能发育
2018-05-04陈剑明刘肖岑徐永健
陈剑明,刘肖岑,徐永健
(宁波大学海洋学院,浙江宁波 315211)
硬骨鱼类的视觉系统较为发达,对摄食、集群等生命活动有着重要的作用[1]。其中,眼球是非常重要的组成器官[1]。研究鱼类视觉系统的发育和结构特征,对阐述其在行为学上的机制及对环境适应性有着重要的意义。
海洋青鳉鱼(Oryziasmelastigma),又名黑点青鳉、印度马达卡(Indian medaka)。因其有对应的淡水青鳉鱼的基因组数据[2],且该鱼具有体型较小、性别差异明显、世代周期短、产卵率高,鱼卵和幼体对环境中各类污染物的干预敏感,盐度适应范围广等特性,目前已被推荐为“一种理想的海洋模式鱼种”[3]。此外,海洋青鳉鱼还具有体色透明易于观察,行为敏捷的特点,也是一种适合研究行为学的良好研究对象。目前针对青鳉鱼视觉器官的形态结构及生理功能的研究尚未见报道。本实验对不同发育阶段的海洋青鳉鱼视觉器官的结构进行组织学研究并探讨其结构组成与功能发育情况,为行为学研究提供理论基础。
当前国内外对鱼类视觉器官的研究主要有以下几个方面:一是鱼类视觉的发育及其与鱼类行为的适应性,如柴毅[4]研究了中华鲟的视觉器官发育及其对摄食的影响,结果表明视觉为中华鲟摄食的第一感觉,对其摄食行为影响不大。Samah 等[5]比较了五种生活在不同海洋深度的硬骨鱼Sardinapilchardus,Gobiusniger,Mullusbarbatusbarbatus和Soleasolea的视网膜结构差异,发现其视网膜结构与其生活在不同水层的习性相适应。二是Fatima等[6]研究了视网膜-顶盖系统,认为视网膜-顶盖系统已经成为研究中枢神经系统发育和再生的一个重要的模型;三是视神经的发育、损伤和恢复也是当前研究较热门的研究课题。如Vera等[7]研究了不同光照强度引起的节细胞、视杆细胞等细胞的凋亡和视网膜各层变化。通过研究调节鱼的视网膜暴露在光照中发生损伤和再生的临界阈值和基本机制,可对养殖过程中使用人工照明的方式进行指导。
1 材料与方法
1.1 材料来源与处理
实验用鱼海洋青鳉鱼来自香港科技大学,经宁波大学培养繁育的子代作为实验材料,实验材料为清洁级。实验地址为宁波大学水产养殖基地(宁波大学东校区)。取2016年8月12日的海洋青鳉鱼1龄成体产出的同一批受精卵,另一培育缸孵化。把刚出膜24 h内的幼体记为0日龄(0 d),出膜0~5日龄的幼体每24 h取样一次,6~30日龄每48~72 h取样一次,30日龄后每10 d取样一次,此外6月龄、9月龄及14月龄个体各取样一次。每次同一批次随机取样12-16尾,均分为两组,其中一组进行暗处理2 h后全部固定;另一组在自然光下,测量有关参数(全长、眼径等),然后再全部固定。30 d内幼体(体长<20 mm),固定整个鱼体,30 d后个体固定整个头部。组织固定液为Bouin氏液,至脱水前24 h再转移至4% 多聚甲醛中。常规石蜡切片,HE染色,显微镜下反复观察,采集图像进行分析。
1.2 测量与计算
1.2.1 绘制图版
根据显微镜下观察并拍摄到的石蜡切片图像,绘制青鳉鱼幼体阶段和成体阶段的眼球示意图(见图1)。
1.2.2 全长、眼径和晶状体直径测量
取待固定的样品鱼置于显微镜下用目微尺测量其体全长(>10 mm的个体用游标卡尺测量)。用Olympus BX53显微镜进行拍照并测量眼球直径(简称眼径)和晶状体直径(简称晶径)。每个实验组均对随机取样的4条青鳉鱼的各项测量指标进行测量统计。
1.2.3 视网膜各层厚度测量
同时,在显微镜下测量眼球中的膜层的层数及各层的厚度。通过有关结果分析判断海洋青鳉鱼的视敏度和光敏度变化[4]。
1.2.4 视网膜运动反应
对比观察海洋青鳉鱼在明暗适应条件下的石蜡切片进行视网膜运动反应分析[4]。
1.3 数据分析
2 结果与分析
2.1 眼球结构
海洋青鳉鱼的眼球结构与其他硬骨鱼相似,包括:角膜(cornea)、虹膜(iris)、脉络膜(chorioidea)、视网膜(retina)、晶状体(lens)、视神经(vision nerve)、银膜(silverskin)、结膜(tunica conjunctive)、环韧带(ligamentum annulare)、水状液(aqueous humor)和玻璃液(vitreous humor)等。各部分出现先后为:
胚胎期能观察到有明显的色素层,说明在感觉器官中眼球是最先分化发育的,初始眼球占头部的比例接近于1/2(图1)。
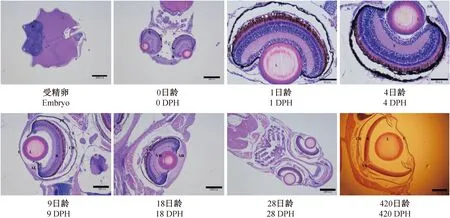
注: C.角膜;I.虹膜;CH.脉络膜;R.视网膜;L.晶状体;VN.视神经;PE.色素上皮层;S.银膜;TC.膜;AL.环韧带;SL.悬韧带;AH.水状液;VH.玻璃液。图1 青鳉鱼眼球的发育Note.C.cornea; I. iris; CH. Chorioidea; R. retina; L. lens; VN. optic nerve; PE. pigment epithelium; S. silverskin; TC. tunica conjunctive; AL. ligamentum annulare; SL. suspensory ligament; AH. aqueous humor; VH. vitreous humor.Fig.1 Development of the medaka eyeball during ontogeny, showing the eyeball structure of medaka at different days post hatching (DPH)
0 d幼体,出现角膜、视网膜和晶状体,但晶状体与视网膜间无空隙,且视网膜厚度大于眼球半径(图1-b)。
1 d幼体,角膜、晶状体、脉络膜和视网膜等都已分化完成。视网膜层厚度接近眼球半径;晶状体呈球状,由无色透明细胞组成;可见脉络膜的三层结构(银膜、血管膜和色素膜);角膜与晶状体间分离,出现水状液;已分化出环韧带,但尚未出现玻璃液。
30 d前,晶状体的大小随眼径一起增长,但晶径与眼径之比基本保持不变(表1)。晶状体与视网膜间的距离越来越远(图1),表明晶状体的前后可调节程度越来越强,即视力不断增强。视网膜层厚度基本保持不变,与眼径的比值在不断下降(图2)。与此同时,晶状体两侧的视网膜在慢慢退化。未观察到悬韧带(suspensor ligament)的存在,但环韧带相当发达。
2.2 视网膜分化
2.2.1 分层情况
海洋青鳉鱼的视网膜由十层结构组成:色素上皮层(PE)、光感受层(PL)、外界膜(OLM)、外核层(ONL)、外网状层(OPL)、内核层(INL)、内网状层(IPL)、神经节细胞层(GCL)、神经纤维层(NFL)、内界膜(ILM)等(图3)。
0 d:视网膜的分化还很不完全,我们可以观察到A区无色素层、感光细胞层、外核层、外网状层等,直接是内核层,同时神经节细胞层占比远大于发育完成的神经节细胞层,可以推测视网膜最先从内核层和神经节细胞层开始分化;B区可观察到还未开始分化的细胞;C区中刚分化的细胞层界限还不清晰,导致在神经节细胞层中出现内网状层(图3-b)。
1 d:色素增多,视细胞层刚开始分化,与外核层无明显分界,无明显外界膜,外网状层已出现,这三层共约占视网膜厚度1/7,而内核层、内网状层、神经节细胞层约占6/7;视细胞纤维也已出现分化,但除外界膜的9层结构已出现外,其余分化还很不完全。视细胞分化基本完成,视锥细胞与视杆细胞均已出现。有明显的外界膜,可观察到完整的视网膜10层结构(图3-a)。
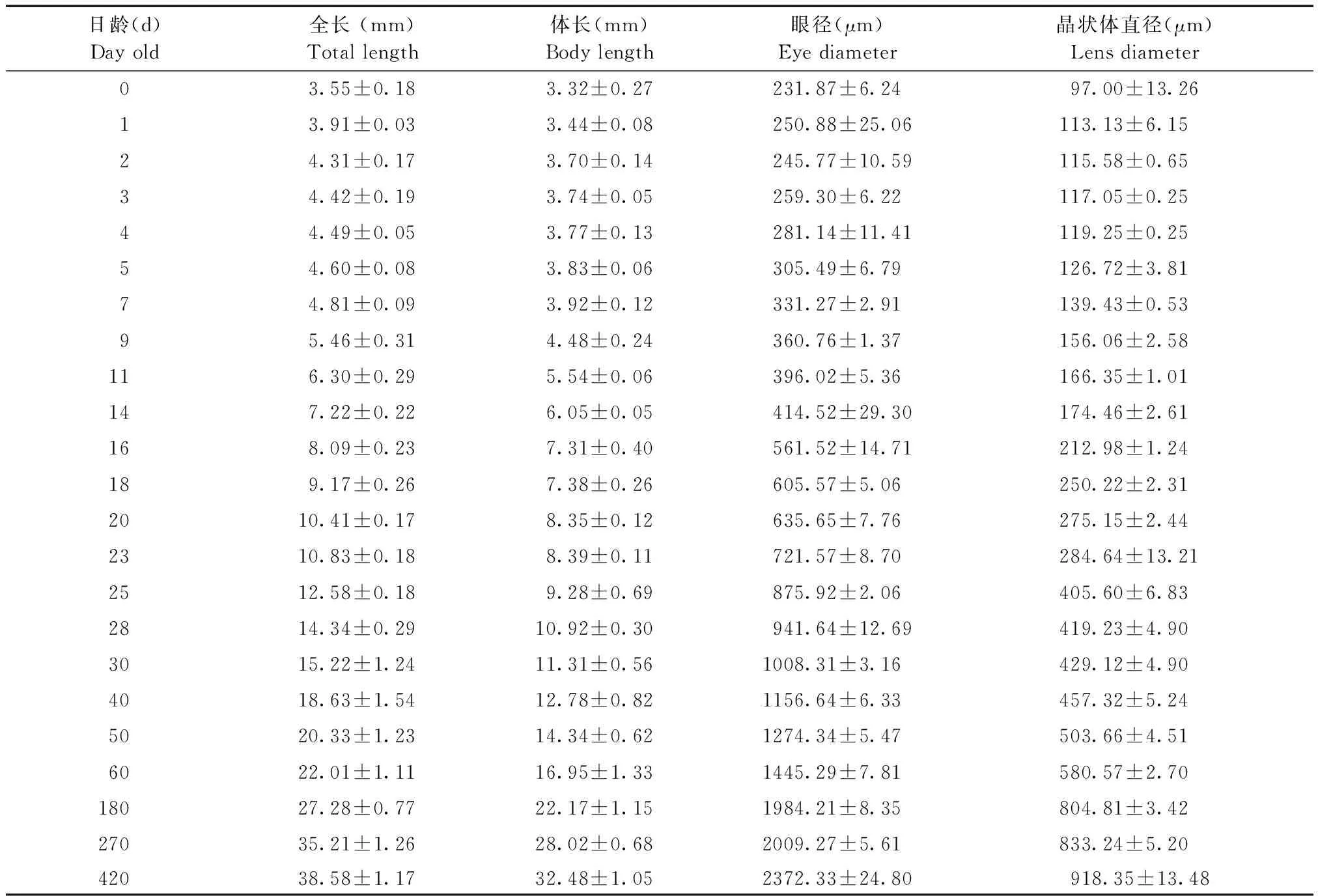
表1 青鳉鱼不同发育阶段形态数据记录表Tab.1 Morphological data of the medaka at different developmental stages
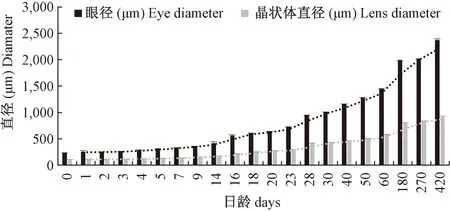
图2 青鳉鱼眼球与晶状体的发育Fig.2 Development of the eye and lens of Medaka
30 d前,视网膜层厚度缓慢增加,但增速远小于眼径的增速。整体而言,视网膜与晶状体间的距离在变远,中间填充了玻璃体,充满玻璃液,用于收集光线,将光线折射到视网膜上。视网膜各层的厚度则发生了各不相同的变化。至2月龄以后,视网膜层的厚度相对于眼径大小,基本上保持了稳定(图3-k、3-l)(图4)。
2.2.2 感光细胞层、外核层及外网状层发育
1 d视网膜的10层结构发育完成后,至60 d感觉细胞层厚度与视网膜层厚度比例基本不变,而后,该比例略有增长(图5)。
外核层主要由视杆细胞的细胞核构成(着色较深),也有视椎细胞存在(着色较浅)。视杆细胞的细胞核排列整齐紧密,呈长柱型;视椎细胞的细胞核在光适应时插入视杆细胞核中间,在暗适应时会后移到外界膜外,细胞核呈卵圆型(图3)。
在发育过程中,外核层与外网状层都会发生一些改变来适应外界变化。从图6看,外核层在幼体发育的前2 d,占视网膜的比例有大幅度减小,之后趋于稳定;至23 d以后至60 d,外核层厚度再次降低,60 d后再次趋于稳定(图6)。外网状层厚度逐渐增加,至6月龄个体达到最大。
2.2.3 内核层与内网状层发育
海洋青鳉鱼的内核层在刚出生时已分化10层以上细胞,其中前3-5层染色较浅、细胞较大、呈圆形的为无长突细胞,中间3~5层染色较深(蓝紫色)、细胞较小、也呈圆形的为双极细胞,后1-2层为水平细胞(图3-a)。随着早期发育完成,青鳉鱼内核层的无长突细胞细胞的细胞层数减少至1~2层细胞,双极细胞与水平细胞层数不变(图3-j)。
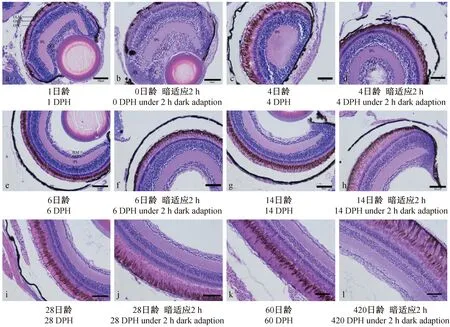
注:PE 色素上皮层;PL 光感受层;OLM 外界膜;ONL 外核层;OPL 外网状层;INL 内核层;IPL 内网状层;GCL 神经节细胞层;NFL 神经纤维层;ILM 内界膜。图3 青鳉鱼视网膜的发育及明暗适应的比较(×400)Note: PE. pigment epithelial layer; PL. photoreceptor layer; OLM. outer limiting membrane; ONL. outer nuclear layer; OPL. outer plexiform layer; INL. inner nuclear layer; IPL. inner plexiform layer; GCL. ganglion cell layer; NFL. nerve fiber layer; ILM. inner limiting membrane.Fig.3 Development and differences between light and dark adaption of retina during medaka ontogeny(×400)
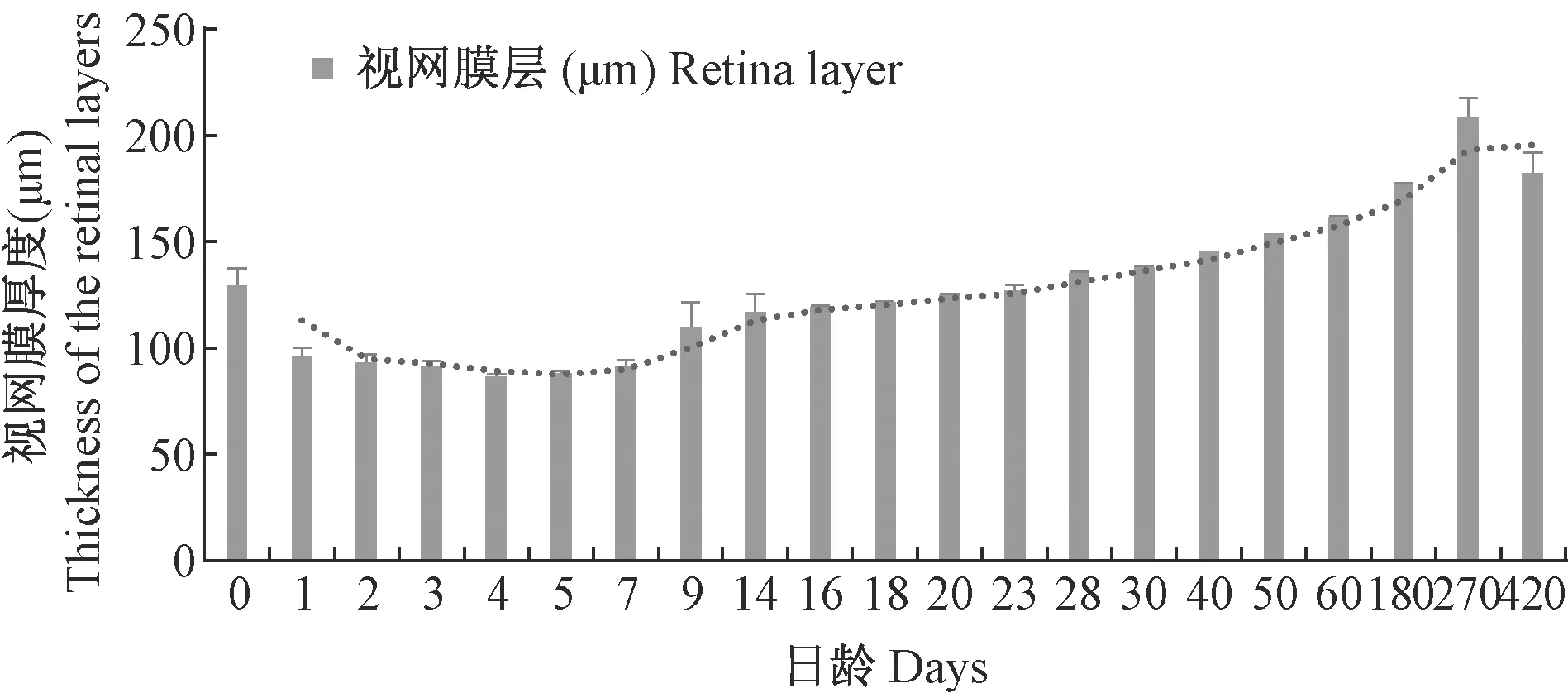
图4 青鳉鱼视网膜层发育情况Fig.4 Development of the retinal layers of medaka
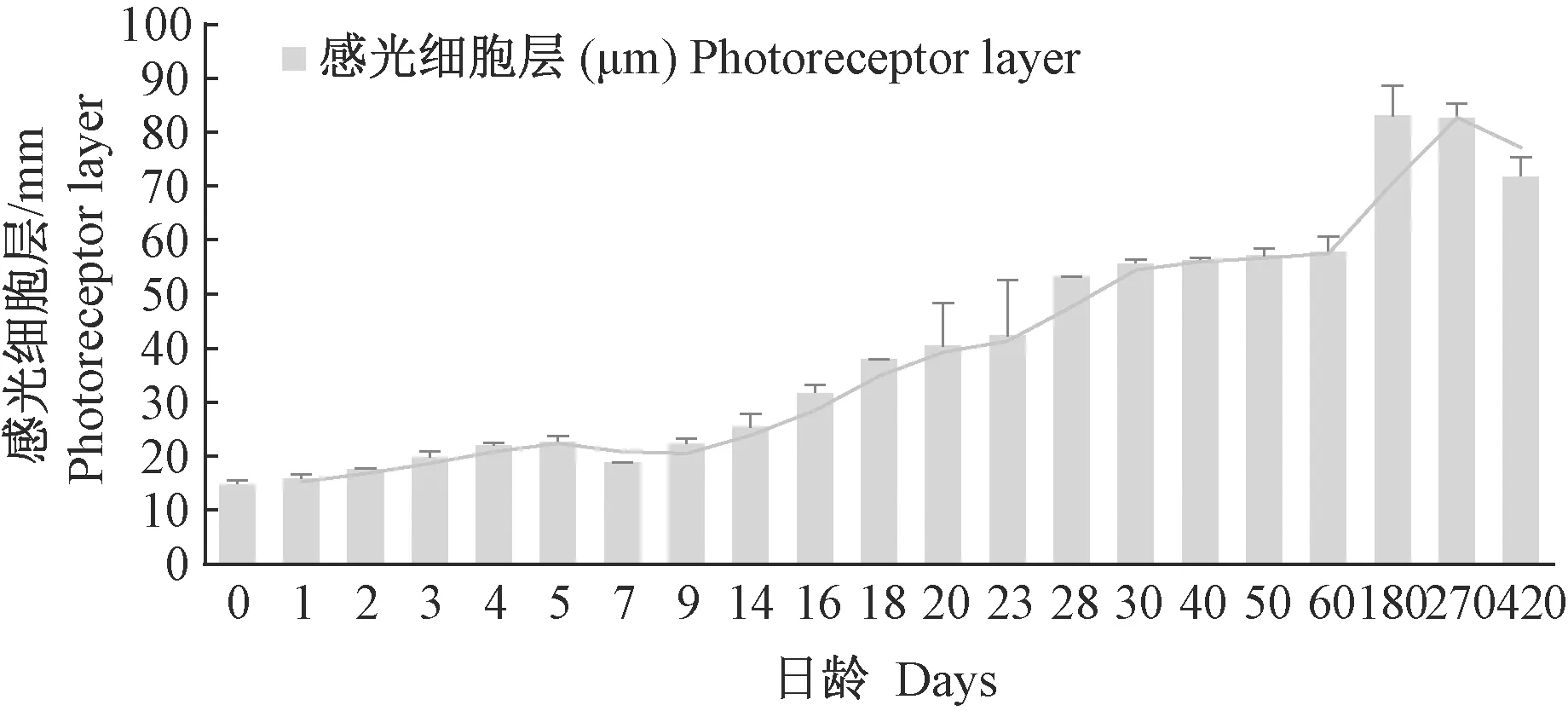
图5 青鳉鱼视网膜感光细胞层的发育Fig.5 Development of the photoreceptor layer of medaka retina

图6 青鳉鱼视网膜外核层和外网状层发育情况Fig.6 Development of the outer nuclear layer and outer plexiform layer of medaka retina
由于视网膜层随着鱼体的生长而生长,而内核层和内网状层在早期就最先发育完成,在后期发育过程中不再发生明显的变化,总体上看,两者占视网膜层的比例在不断减少(表2)。从图6看,刚孵化的青鳉鱼的内核层和内网状层厚度有所下降,4 d后两者的厚度上下波动;至14 d后,内核层的厚度缓慢降低(图7)。
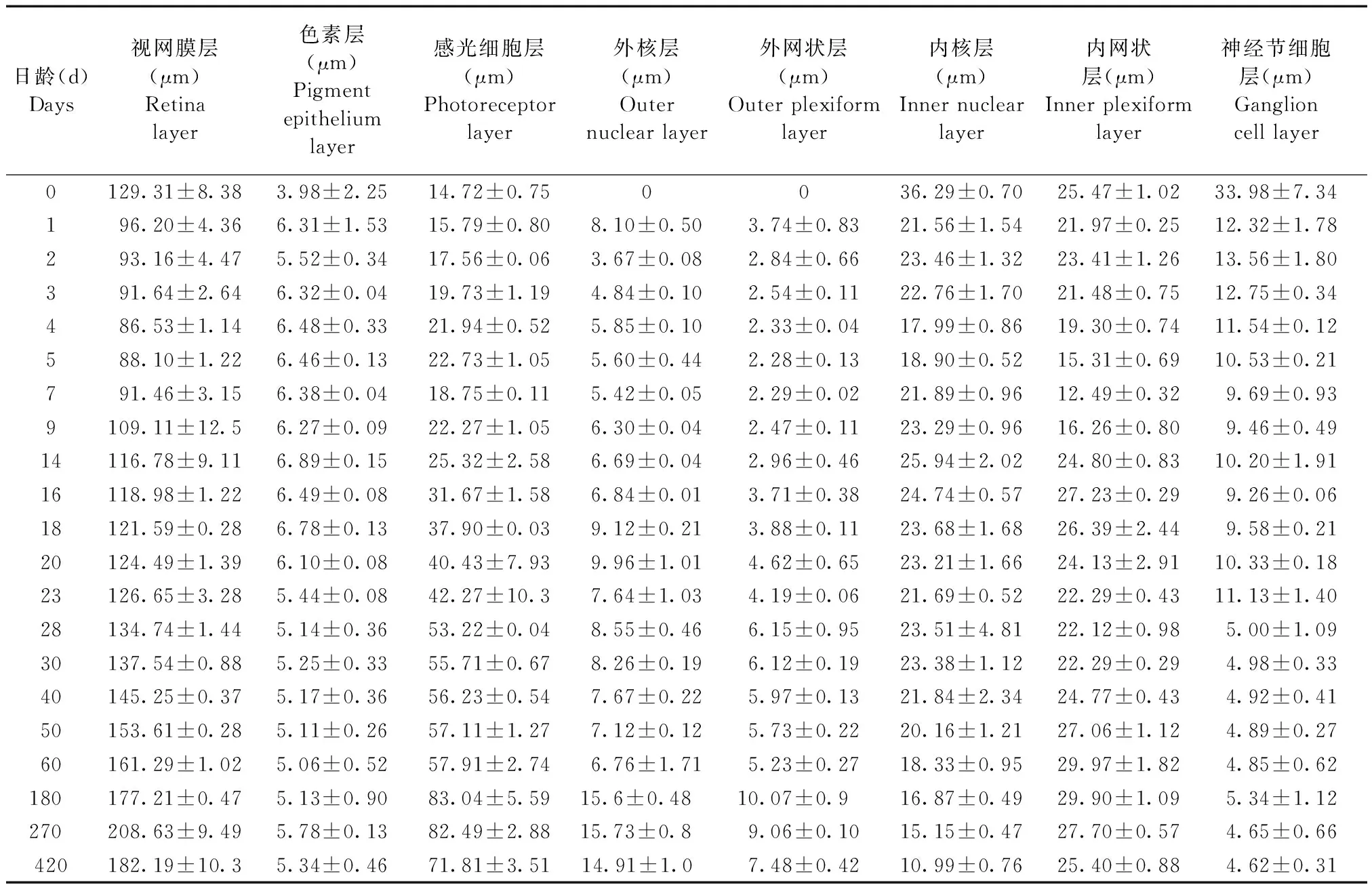
表2 青鳉鱼不同发育阶段视网膜各层变化统计表Tab.2 Changes of the retinal layers at different developmental stages of medaka
2.2.4 神经节细胞层的发育
神经节细胞的细胞核呈圆形,排列较为紧密,染色也较深,呈蓝紫色(图3)。海洋青鳉鱼初孵12 h时,神经节细胞层的层数不规则,厚的部位多达到11层,薄的部位仅为6层(图3-a);2~4 d,神经节细胞层的层数已均等,为6~8层;5 d,减少至3层(图3-c);15 d,减少至2层;28 d,向一层细胞过渡(图3-g),60 d后,为1层细胞(图3-i)。神经节细胞层的厚度也随其细胞层数减小而下降(图8)。
青鳉鱼视网膜视细胞纤维层在靠近视神经部分细胞层数增加,靠近角膜部分只有一层细胞。
2.3 视网膜运动反应
对比发育过程中对比青鳉鱼发育过程中明适应与暗适应状态下的视网膜变化(图3),可以发现:
无论暗适应还是明适应的条件下,0至2 d青鳉鱼的色素层逐渐增多,均无位移发生(图3-a、3-b),12 h内只分化出少量视杆细胞;1日龄时视杆细胞与视椎细胞均出现,至2 d视椎细胞无前移现象,说明此时仔鱼的视觉很弱。
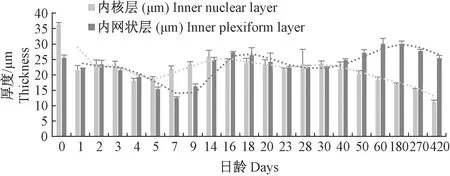
图7 青鳉鱼视网膜内核层和内网状层发育情况Fig.7 Development of the inner nuclear layer and inner plexiform layer of medaka retina
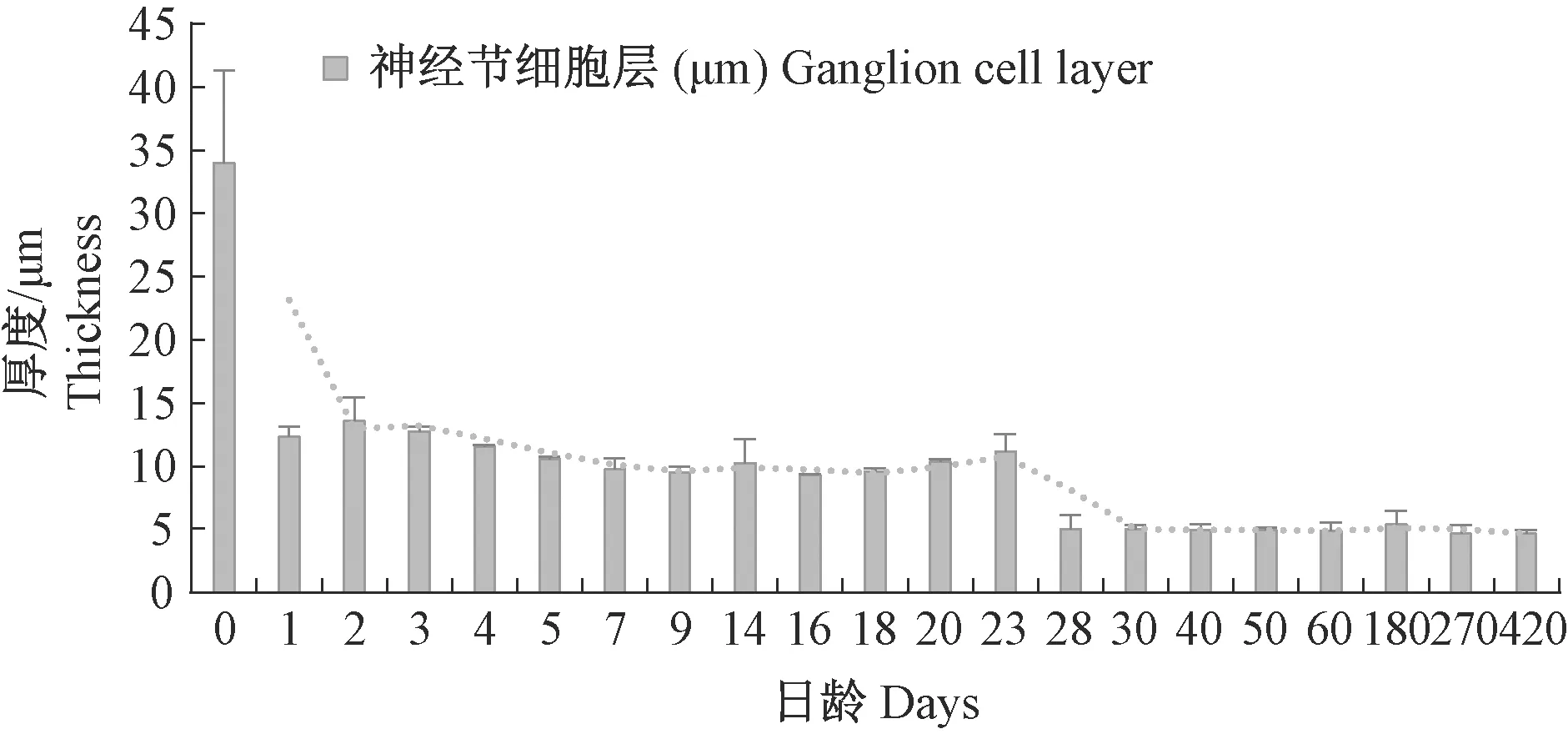
图8 青鳉鱼视网膜神经节细胞层发育情况Fig.8 Development of the ganglion cell layer of Medaka retina
4 d,暗适应和明适应鱼体的色素层均有相同的前移现象。明适应下,视椎细胞存在于视杆细胞间,而暗适应时,视椎细胞与外界膜接触,有微小的前移现象。说明有可能已经出现明视觉,而暗视觉处于形成阶段(图3-c、3-d)。
6 d,明适应的青鳉鱼的色素层前移包裹视杆细胞(防光散射),视椎细胞也前移,穿过外界膜,进入外核层区。而暗适应2 h后的青鳉鱼色素层后移,回到色素细胞下方,视杆细胞暴露。因此,推测5 d青鳉鱼已具有明视觉与暗视觉能力(图8-e、8-f)。
14~15 d,青鳉鱼的视网膜运动现象非常明显,表明已有很强的明视觉与暗视觉能力(图3-g、3-h)。
3 讨论
海洋青鳉鱼的视觉器官结构完整、发育迅速。在胚胎期即可观察到眼球雏形,至1日龄,可以在显微镜下观察到其眼球前后转动,可以观察到晶状体在眼球内的转动;此时,海洋青鳉鱼迅速分化出了角膜、晶状体、视网膜、脉络膜等,其中视网膜包围着晶状体,晶状体两侧的两部分与虹膜整个内侧相连为盲区,后面部分与脉络膜的血管膜相连;但感光细胞发育不完全,也没有水状液和玻璃液收集折射光线,推测已经存在较弱的视觉。1日龄后,眼球结构基本完整,盲区的视网膜开始退化,至28日龄盲区的视网膜厚度只有虹膜的1/2,60日龄时退化至1/3,9月龄时盲区已经完全退化,而晶状体与视网膜的距离随着眼球增大而不断增大,视觉能力逐渐增强。柴毅(2006年)[4]认为中华鲟仔鱼开口饵料的捕食主要靠视觉器官,随着其他感觉器官的发育,视觉退化。而青鳉鱼眼球发育早,且视觉器官没有退化现象,视觉能力不断增强,可以判断视觉是其捕食、避害的重要器官。
梁旭方等[8]的研究表明光感受系统发达的中上层鱼类,其内核层发育成典型的4层水平细胞,且分化出明显的水平细胞、双极细胞和无长突细胞,其中水平细胞与双极细胞位于内核层的外侧。马爱军等[9]研究表明半滑舌鳎的层数少,且水平细胞、双极细胞和无长突细胞分化不明显是光感受系统不发达的底栖鱼类的类型,且视网膜的光感受细胞排列不规则,无镶嵌结构,表明其视敏度与光敏度均不发达,与其底栖生活和白天不摄食的行为相适应。相比之下,海洋青鳉鱼为中上层鱼类[10],一般认为中上层鱼类都有发达的光感受系统[11],我们观察到其内核层分化出明显的水平细胞、双极细胞和无长突细胞,其中水平细胞与双极细胞位于内核层的外侧;而底栖鱼类的内核层层数少,且水平细胞、双极细胞和无长突细胞分化不明显是光感受系统不发达的底栖鱼类的类型。本研究也发现:海洋青鳉鱼的水平细胞、双极细胞和无长突细胞分化很明显,表明其是中上层鱼类,但其水平细胞最终只发育成1层。这一现象与牙鲆的光感觉系统类似,牙鲆的内核层也分化明显,但水平细胞仅有1-2层,这与其营底栖生活却白天摄食为主的习性相适应[12]。因此,海洋青鳉鱼和牙鲆的光感受系统发达水平一致,习性也应相似,这种视觉结构的变化与其从浮游到底栖的生态迁移及捕食方式的变化相适应[13]。
同时,本研究还发现青鳉鱼感光细胞层的单锥细胞和双锥细胞形成的典型镶嵌结构,表明鱼眼对运动具有较强的敏感性。该镶嵌结构表明鱼眼有较强的视敏度,即对运动具有较强的敏感性[8],可以在较弱光场条件下提高对视觉对象的分辨能力[14]。海洋青鳉鱼的视锥细胞,椎体细长,排列规则,呈典型的镶嵌结构,因此推断这可以增强运动的敏感性,且光敏感性和视敏度强,这符合海洋青鳉鱼以白天摄食为主,且运动敏捷,而在暗环境中几乎不摄食不运动的习性。早期有研究认为:视觉对鱼类摄食的影响主要具有寻找和发现,辨认和选择,摄食时方向和姿势的调整等作用[15]。因此,可以推断海洋青鳉鱼的视觉在其捕食与游泳行为中具有重要的意义。
此外,生物的视网膜结构特征是与其生活习性相一致的,如魏开建(1996)[15]发现鳜鱼视觉结构的变化与其从浮游到底栖的生态迁移及捕食方式的变化相适应;江剑平等(2001)[16]也发现虎纹蛙视网膜结构特征与其生活于田间、山涧等弱光环境和捕食习性是相适应的。我们观察发现,海洋青鳉鱼的视网膜在1日龄已经是10层结构,至5日龄出现视网膜运动,23日龄开始变薄,与晶状体间距离越来越大,这既有利于调整晶状体的前后位置,便于随时观察较近和较远距离的物体,同时视网膜上可投射光的面积也大大增加,表明其视觉功能随着发育越来越完善。海洋青鳉鱼发育至第5天,出现视网膜运动现象,至14~15 d,视网膜运动现象很明显。视网膜运动反应是适应明暗环境感光细胞发生的位置迁移,是视觉能力强弱的表现[17-18]。表明该时期,海洋青鳉鱼具有了良好的明视觉与暗视觉的调节能力。这就有利于其摄食、游泳、避敌等生命活动,这也符合其为中上层鱼类对光线明暗较为敏感的适应特征[12-14]。而在视网膜各层的发育过程中,观察到在4~5日龄,14~16日龄,23~28日龄有较为显著的变化,且视网膜层厚度占眼径比,内核层和神经节细胞层占视网膜层厚度比等随着发育均有减小现象。尤其是14~28日龄,视网膜结构发生较大变化,而该阶段为海洋青鳉鱼由浮游变为底层活动的过程,这说明青鳉鱼的视网膜结构的发育与其生活习性变化相适应。
综上所述,本实验研究海洋青鳉鱼的视觉发育为下一步研究海洋青鳉鱼的行为学提供了理论依据。作为新的模式生物,对海洋青鳉鱼的研究可对其他物种的研究提供了较好的实验方法,以期获得更完善、更丰富的实验数据和科学结论。
参考文献(References)
[1] 苏锦祥. 鱼类学与海水鱼类养殖 第二版 [M]. 北京.中国农业出版. 1993.
Su JX. Ichthyology and Marine Fish Cultivation. 2nd Edition. [M]. Beijing: China Agriculture Press, 1992
[2] Kong R, Giesy JP, Wu R, et al. Development of a marine fish model for studying in vivo molecular responses in ecotoxicology [J]. Aquatic Toxicol, 2008, 86(2): 131~141.
[3] Wu XL, Huang QS, Fang C, et al.Oryziasmelastigma: A new promising model organism for marine ecotoxicology [J]. Asian J Ecotoxicol, 2012, 7(4): 345-353.
[4] Chai Y. Development of sense organs and the behavior function during Chinese sturgeon (AcipenserSinensis) Ontodeny [D]. Thesis for Doctor of Science. Huazhong Agricultural University, 2006.
[5] Darwish ST, Mohalal ME, Helal MM, et al. Structural and functional analysis of ocular regions of five marine teleost fishes (Hippocampus hippocampus, Sardina pilchardus, Gobius niger, Mullus barbatus & Solea solea) [J]. Egypt J Basic Appl Sci, 2015, 2(3): 159-166.
[6] Fatima A, Martin PM. Fish vision: size selectivity in the zebrafish retinotectal pathway [J]. Curr Biol, 2014, 24(21): 1048-1050.
[7] Vera LM, Migaud H. Continuous high light intensity can induce retinal degeneration in Atlantic salmon, Atlantic cod and European sea bass [J]. Aquaculture, 2009, 296: 151-158.
[8] Liang XF. Visual characteristics of mandarin fish (Siniperca chuatsi) in relation to its feeding habit: II-general properties of the retina [J]. Acta Hydrobiol Sin, 1998,18(4): 376-377.
[9] Ma AJ, Wang XA, Zhuang ZM, et al. Structure of retina and visual characteristics of the half-smooth tongue-soleCynoglossussemilaevisGünter [J]. Acta Zoolog Sin, 2007, 53(2): 354-363.
[10] 陈漪, 王晓杰, 冉皓宇, 等. 海水模式种青鳉鱼(Oryziasmelastigma) 的胚胎发育观察 [J]. 海洋与湖沼, 2016, 47(1): 71-82.
Chen Y, Wang XJ, Ran HY, et al. Developmental stages of a marine model fish——MedakaOryziasMelastigma[J]. Oceanol Limnol Sin, 2016, 47(1): 71-82.
[11] Liang XF, He DR. Sensory basis in the feeding behivior of fishes [J]. Acta Hydrobiol Sin, 2007, 53(2): 354-363.
[12] Lin LM, Li YY, Wang RJ, et al. A study on feeding rhythm ofParalichthysolivaceusduring early developmental stage [J]. J Fisheries Chin, 2006, 30(3): 329-334.
[13] He DR, Xu YG. Comparison on retinal structure of five species of marine fishes [J]. J Oceanogr Taiwan Strait, 1993, 12(4): 342-350.
[14] Shan BR, He DR. The relation between feeding and vision development of black porgy [J]. J Oceanogr Taiwan Strait,1995,14(2): 169-173.
[15] Wei KJ, Zhang GR, Zhang HM. Studies on the phototactic characteristics of mandarin fish (Sinipercachuatsi) during different development stages [J]. J Huazhong Agr Univ, 2001, 20(2): 164-168.
[16] Jiang JP, Cheng WL, Yu W, et al. Studies on retinal structure ofRanaRugulosaand its environmental adaptation [J]. Chin J Appl Environm Biol, 2001, 7(1): 51-55.
[17] 李霞. 水产动物组织胚胎学 [M]. 北京. 中国农业出版. 2005.
Li X. Histology and Embryology of Farming Aquatic Animals [M]. Beijing: China Agriculture Press. 2005
[18] Xi D. Growth pattern and cannibalism in Black Pockfish (Sebastesschlegelii) during early development stage [D]. Thesis for Master of Science. Ocean University of China, 2014.
