Toll样受体2基因敲除对高脂饮食诱导的肥胖小鼠脂肪组织凋亡的影响
2018-04-17张浩强张素萍沈雁飞
张浩强 张素萍 沈雁飞 宋 冰
(锦州医科大学附属第一医院内分泌科,辽宁 锦州 121000)
肥胖机体存在低水平的内毒素血症〔1〕,而内毒素作为革兰阳性菌产物可以激活Toll样受体(TLRs)通路引起机体炎症反应〔2〕。脂肪组织的炎症反应和细胞的凋亡相关〔3〕,肥胖小鼠脂肪组织细胞凋亡增加,且膳食结构的质和量可以影响脂肪组织凋亡〔4〕。TLR2属于TLRs家族中的一员,是与固有免疫相关的一种模式识别受体,能够识别多种病原相关分子模式,与糖脂代谢有关〔2〕。TLR2缺乏可减轻高脂饮食诱导的脂肪组织炎症反应〔5〕,因此猜想,TLR2基因可能调节高脂饮食诱导的肥胖小鼠脂肪组织的细胞凋亡。本实验采用TLR2基因敲除的小鼠,并给予高脂饮食诱导肥胖,观察脂肪组织凋亡相关蛋白的变化。
1 材料与方法
1.1动物分组与处理SPF级4周龄C57BL/6J雄性小鼠24只(SCXK〔京〕2009-0015,华阜康公司,北京)和TLR2基因敲除小鼠24只(022507,Jackson 实验室,美国),体质量(14±2)g,随机分为正常对照组(NC,8只)、肥胖组(OB,16只)、TLR2基因敲除组(TK,8只)和TLR2基因敲除肥胖组(TO,16只)。自由饮水、摄食,动物房内12 h昼夜循环,室温(22±2)℃。专用高能量饲料(华阜康公司,北京)喂养OB组和TK组小鼠,小鼠全价营养颗粒基础饲料(华阜康公司,北京)喂养NC组和TK组小鼠,16 w后OB组小鼠体重大于NC组小鼠上限者及TO组小鼠体重大于TK组小鼠上限者随机各选取8只用于实验,其余剔除。
1.2生化指标检测16 w后,各组小鼠禁食6 h,心脏取血,留取血清测定空腹血糖(FPG),三酰甘油(TG),总胆固醇(TC),高密度脂蛋白(HDL)和低密度脂蛋白(LDL)。
1.3caspase3活性检测小鼠处死后,速取部分腹腔脂肪组织,置入-80℃冰箱中备用。提取组织总蛋白,BCA蛋白定量试剂盒(博奥森公司,北京)测总浓度,并调整各蛋白浓度2 mg/ml,-80℃冰箱保存备用。采用caspase3分光光度法检测试剂盒(万类,沈阳)检测caspase3活性。
1.4Western印迹取材及蛋白定量同1.3。十二烷基硫酸钠-聚丙烯酰胺凝胶电(SDS-PAGE):分离胶(12%)及浓缩胶(5%),每孔20 μl样本及小分子Marker(赛默飞,上海)上样进行电泳。 采用湿转法转蛋白至聚偏氟乙烯(PVDF)膜,脱脂奶粉封闭,bcl2和bax多克隆抗体(万类,沈阳)4℃过夜,过氧化物酶标记的山羊抗兔二抗(ABSCI,美国)孵育2 h。用GISt020凝胶图像分析仪及成像。
1.5荧光实时定量PCR脂肪组织总RNA采用Trizol抽提,包含反转录酶的RNeasy Plus Mini试剂盒(Bioneer,韩国)将mRNA反转录为cDNA,QIAGEN Fast Cycling PCR 试剂盒(Takara生物科技,大连)进行扩增。引物由上海生工设计并合成,bcl2:正义5′-CCCCTCGCATCTTCTCCTTCC-3′,反义5′-CCACCACCTCCTTGAGAAGTCC-3′;bax:正义5′-CCAGGATGCGTCCACCAA-3′,反义5′-CACCAACGGGAGAAGATGAAACG-3′;GAPDH:正义5′-TTGTCAAGCTCATTTCCTGGTATG-3′,反义5′-GGATAGGGCCTCTCTTGCTCA-3′,所得数据以GAPDH mRNA作为参照。
1.6统计学方法采用SPSS20.0软件行one way NOVA方差分析,两两比较用LSD法和Tamhane法。
2 结 果
2.1高脂饮食和TLR2基因敲除对小鼠血糖、血脂的影响与NC组〔(6.78±0.43)、(2.64±0.06)、(0.26±0.02)、(1.46±0.02)mmol/L〕相比,OB组FPG、TG、TC和LDL水平〔(9.79±0.32)、(3.03±0.07)、(0.37±0.03)、(1.65±0.07)mmol/L〕明显升高,HDL水平〔(1.05±0.02)mmol/L〕明显降低(P<0.05)。与OB组相比,TO组小鼠FPG〔(6.88±0.33)mmol/L〕、TG〔(2.36±0.06)mmol/L〕、TC〔(0.12±0.02)mmol/L〕、HDL〔(0.94±0.02)mmol/L〕和LDL水平〔(1.27±0.04)mmol/L〕明显降低(P<0.05)。TK组FPG、TG、TC、HDL、LDL水平分别为(6.37±0.46)、(2.30±0.06)、(0.08±0.01)、(0.98±0.03)、(1.19±0.03)mmol/L。
2.2高脂饮食和TLR2基因敲除对小鼠脂肪组织caspase3活性的影响OB组(35.37±2.96)与NC组(23.10±3.15)相比,小鼠脂肪组织caspase3活性明显升高(P<0.05),TO组(23.95±3.53)与OB组相比,小鼠脂肪组织的caspase3活性降低(P<0.05)。TK组Caspase3活性为18.28±3.93。
2.3高脂饮食和TLR2基因敲除对小鼠脂肪组织bcl2和bax蛋白的影响与NC组(0.56±0.05,0.33±0.23)相比,OB组脂肪组织bcl2蛋白量减少(0.24±0.07,P<0.05),bax蛋白量增加(0.62±0.10,P<0.05)。与OB组相比,TO组脂肪组织bcl2蛋白量增加(0.52±0.04,P<0.05),bax蛋白量减少(0.36±0.03,P<0.05)。与NC组(0.60±0.09)相比,OB组bax蛋白与bcl2蛋白比值(2.63±0.31,P<0.05)增加;与OB组相比,TO组bax蛋白与bcl2蛋白比值降低(0.70±0.02,P<0.05)。TK组bax蛋白量0.24±0.04,bcl2 0.73±0.07,bax/bcl2 0.33±0.10。见图1。
2.4高脂饮食和TLR2基因敲除对小鼠脂肪组织bcl2和bax mRNA的影响与NC组(0.71±0.02,0.35±0.02)相比,OB组脂肪组织bcl2 mRNA减少(0.22±0.03,P<0.05),bax mRNA增加(0.94±0.02,P<0.05)。与OB组相比,TO组脂肪组织bcl2 mRNA增加(0.61±0.05,P<0.05),bax mRNA减少(0.54±0.02,P<0.05)。与NC组(0.49±0.02)相比,OB组bax mRNA与bcl2 mRNA比值增加(4.43±0.67,P<0.05);与OB组相比,TO组bax mRNA与bcl2 mRNA比值降低(0.90±0.09,P<0.05)。TK组bcl2 mRNA为0.90±0.03,bax mRNA为0.24±0.02,bax/bcl2 mRNA为0.26±0.02。
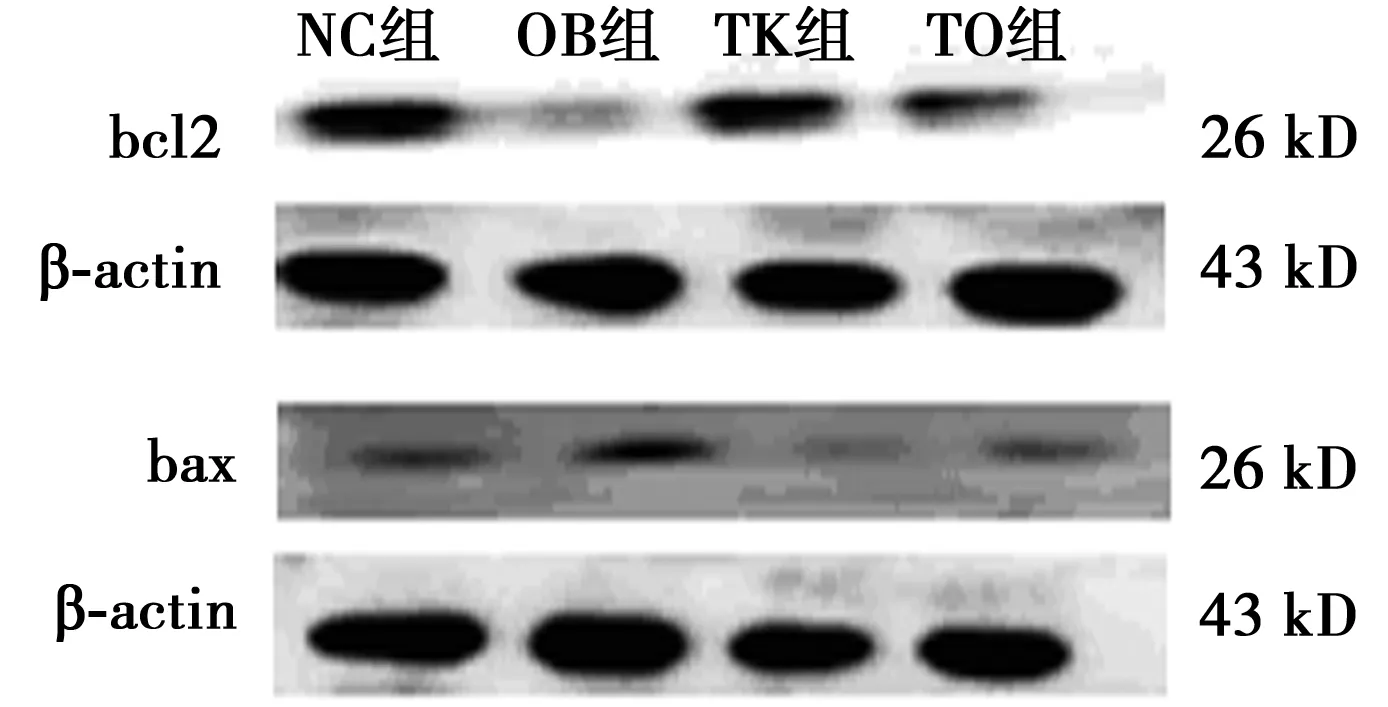
图1 小鼠脂肪组织bax和bcl2蛋白表达
3 讨 论
TLR2与固有免疫相关,是模式识别受体家族的重要一员,能够识别包括脂肪酸在内的病原相关分子模式〔6〕。有研究认为膳食结构改变可以影响脂肪组织凋亡相关蛋白的表达,且肥胖小鼠脂肪组织细胞凋亡增加〔4〕,TLR2敲除对肥胖小鼠脂肪组织细胞凋亡的影响尚不明确。caspase3和细胞凋亡相关,常作为衡量细胞凋亡程度的指标〔7,8〕,本研究结果说明TLR2基因敲除可能减轻了高脂饮食诱导的肥胖小鼠脂肪组织细胞凋亡。bcl2家族蛋白在细胞凋亡中的作用分为两类:一类是以bcl2为代表的抗凋亡蛋白,一类是以bax为代表的促凋亡蛋白。bcl2和Bax可以形成异源二聚体,阻断细胞色素(Cyt)-C的释放,继而抑制caspase-3蛋白活化,有效抑制细胞凋亡〔9〕bcl2和bax的比值增高提示抑制细胞凋亡的作用增强,反之,提示促进细胞凋亡作用增强,因此bax/bcl2蛋白比值决定了细胞的生存和死亡〔10,11〕。本研究提示,TLR2基因敲除可能减轻了高脂饮食诱导的肥胖小鼠脂肪组织细胞凋亡,并且这可能与其发挥降脂作用相关。
1Kim KA,Gu W,Lee IA,etal.High fat diet-induced gut microbiota exacerbates inflammation and obesity in mice via the TLR4 signaling pathway〔J〕.PLoS One,2012;7(10):e47713.
2Kawasaki T,Kawai T.Toll-like receptor signaling pathways〔J〕.Front Immunol,2014;5:461.
3Mocanu AO,Mulya A,Huang H,etal.Effect of Roux-en-Y gastric bypass on the NLRP3 inflammasome in adipose tissue from obese rats〔J〕.PLoS One,2015;10(10):e0139764.
4Camargo A,Rangel-Zúiga OA,Alcalá-Díaz J,etal.Dietary fat may modulate adipose tissue homeostasis through the processes of autophagy and apoptosis〔J〕.Eur J Nutr,2016.〔Epub ahead of print〕.
5Davis JE,Braucher DR,Walker-Daniels J,etal.Absence of Tlr2 protects against high-fat diet-induced inflammation and results in greater insulin-stimulated glucose transport in cultured adipocytes〔J〕.J Nutr Biochem,2011;22(11):136-41.
6Xu Y,Wang LM,He J,etal.Prevalence and control of diabetes in Chinese adults〔J〕.JAMA,2013;310(9):948-58.
7Bernardi P,Rasola A.Calcium and cell death:the mitochondrial connection〔J〕.Subcell Biochem,2007;45(1):481-506.
8Lin M,Li L,Li LP,etal.The protective effect of baicalin against renal ischemia-reperfusion injury through inhibition of inflammation and apoptosis〔J〕.BMC Complement Altern Med,2014;14(1):19.
9Lee Y,Gustafsson AB.Role of apoptosis in cardiovascular disease〔J〕.Apoptosis,2009;14(4):536-48.
10Bouillet P,Purton JF,Godfrey DI,etal.BH3-only Bcl-2 familly member Bim is required for apoptosis of autoreactive thymocytes〔J〕.Nature,2002;415(6874):922-6.
11Antonsson B.Mitochondria and the Bcl-2 family proteins in apoptosis signaling pathways〔J〕.Mol Cell Biochem,2004;256(1/2):141-55.
