蛋白质对黄酒品质影响的研究进展
2018-04-12齐小慧孙军勇谢广发陆健
齐小慧,孙军勇,谢广发,陆健*
1 (江南大学,工业生物技术教育部重点实验室,江苏 无锡,214122) 2(江南大学,粮食发酵工艺与技术国家工程实验室,江苏 无锡,214122) 3(江南大学 生物工程学院,江苏 无锡,214122)4(中国绍兴黄酒集团有限公司 国家黄酒工程技术研究中心,浙江 绍兴,312000)
根据国家标准GB/T 13662—2008《黄酒》[1]的定义,黄酒是指以稻米、黍米等为主要原料,经加曲、酵母等糖化发酵剂酿制而成的发酵酒。黄酒中含有丰富的氨基酸、无机盐、微量元素、维生素和多种活性物质,不仅营养丰富,而且长期以来被认为是保健养生品,还可用作中药药引服用[2]。实验证明[3],饮用黄酒有助于排铅、增强记忆力和免疫力、抗氧化、抗衰老、预防骨质疏松等。蛋白质在黄酒中含量比较丰富,为各酿造酒之首[4-6]。绍兴花雕酒蛋白质含量为5.78 g/L,绍兴加饭酒总蛋白质含量高达16 g/L,是啤酒的4倍,红葡萄酒的16倍[7-8]。黄酒中的蛋白质作为黄酒重要的组成成分,对黄酒的营养价值、风味及保健功能等都有较显著影响,而蛋白质对黄酒的非生物稳定性的影响最为显著。
黄酒作为中国最古老的酒种,具有数千年的发展历史,曾是全国性的饮料酒,但由于各种原因,黄酒目前的发展已经落后于整个酒业的发展速度,年产量与产值远低于白酒,且与葡萄酒相比,虽然其年产量较高,年产值却低于葡萄酒[9]。近年来,黄酒因其营养、保健等功效被熟知,逐渐被重视,加强对黄酒成分的研究将有助于黄酒产业的发展。本文从黄酒蛋白的角度出发,综述了黄酒蛋白的来源、含量、分布以及其对黄酒品质的影响,提出目前存在的问题及今后的研究方向。
1 黄酒酒体的蛋白质
近年来,对黄酒蛋白质的研究大多是围绕着浑浊蛋白进行的,对于酒体蛋白的研究相对较少。通过分析不同类型及陈酿年份的黄酒发现[4,7,10],黄酒中低分子蛋白的含量最高,高分子蛋白的含量最少,且随着陈酿年份的增加而发生改变:当年新酒中蛋白质的分子质量以10 kDa左右为主,而随着陈酿年份的增加,低分子质量蛋白的含量变化不明显,高分子量蛋白的含量明显下降。
目前对于黄酒酒体蛋白的种类、数量及来源的研究还未开展,一些研究者认为[8],黄酒中的蛋白质主要来源于大米和麦曲,还有少量的微生物蛋白和酶蛋白。通过合适的方法分离出酒体蛋白,并采用双向电泳技术及基质辅助激光解析电离飞行时间串联质谱法等方法,可以鉴定黄酒酒体蛋白的种类、数量及来源。
1.1 麦曲来源的蛋白质
小麦中含有大量的可溶性蛋白,在制曲过程中由于米曲霉等微生物的存在,一些小麦蛋白质会被降解;在对浑浊蛋白的研究中鉴定出小麦来源的浑浊蛋白,说明也有一些小麦蛋白未被降解,如二聚-α-淀粉酶抑制剂等[11-12]。
张波等[13]对未接种熟小麦、熟麦曲及制曲过程中4个不同时期(即孢子萌发期、菌丝生长期、菌丝繁殖期和孢子着生期)的麦曲中宏蛋白质组进行分析,结果表明,未接种熟小麦中的蛋白为原料小麦蛋白,到菌丝繁殖期原料小麦蛋白基本被米曲霉分泌的蛋白酶降解。ODA等[14]蛋白质组学分析也表明,小麦蛋白在培养24 h时几乎被米曲霉分泌的蛋白酶降解。有研究者认为[15],绍兴黄酒成品麦曲中的小麦蛋白绝大部分在制曲过程中被降解,因此麦曲中的可溶性蛋白大部分为微生物分泌的胞外酶。但孔令琼[16]对黄酒麦曲中的宏蛋白质组进行分析时却发现,鉴定出的124个蛋白点中非生物来源的蛋白点共有113个,其中有100个蛋白点来源于小麦,包括22种蛋白;6个蛋白点来源于水稻,包括3种蛋白;而微生物来源的蛋白点只有11个;鉴定出的非生物来源的蛋白质主要是酶抑制剂类,例如α-淀粉酶抑制剂等,另外还有水解酶类以及少量其他酶。造成这种结果差异的原因可能是选取了制曲过程不同时期的麦曲作为实验样品。
1.2 大米来源的蛋白质
传统黄酒酿造多选用糯米为原料,糯米中含有丰富的淀粉、蛋白质、脂肪、维生素等[17]。与原料小麦中的蛋白质类似,原料糯米中的蛋白质经过浸泡、蒸煮和发酵后,大部分已经被蛋白酶分解为肽和氨基酸,还有少量蛋白质未被分解保留在酒液中,例如类燕麦蛋白和β-淀粉酶[11,18]。经研究发现[18],糯米中总蛋白含量为8.10%~10.30%之间,可溶性蛋白含量为0.15 ~0.49 mg/L,糯米中蛋白质的含量在很大程度上影响黄酒中游离氨基酸的含量。代钰等[19]采用稀碱法提取糯米蛋白,确定了最佳提取工艺:碱液浓度0.05 mol/L,温度45.68 ℃,料水比8∶1,时间94.56 min,但未对糯米蛋白的种类进行鉴定。
1.3 酵母来源的蛋白质
有关葡萄酒的研究表明[20],在葡萄酒的储酒阶段,自溶和死亡的酵母体内的蛋白质和蛋白酶会释放到酒液中,对成品葡萄酒的蛋白稳定性产生影响。黄酒中也有可能存在类似的问题,因为黄酒有长达4个月的后酵期,酵母也可能会自溶、死亡,其体内的蛋白质也会释放到酒液中,但此推测是否正确还需对黄酒酒体蛋白及其来源做进一步的研究。
2 蛋白质对黄酒品质的影响
2.1 蛋白质对黄酒营养成分的影响
蛋白质在黄酒中主要以氨基酸和肽的形式存在,人体容易吸收利用。氨基酸是一种重要的营养成分,在黄酒中的含量极其丰富,种类多达21种,其中包括8种必需氨基酸,且在各酿造酒中必需氨基酸的含量最丰富、种类最齐全[8]。绍兴加饭酒中含有4.3 g/L左右的游离氨基酸,其中必需氨基酸约占34.9%,半必需氨基酸约占27.9%;绍兴加饭酒中的总氨基酸含量是日本清酒的1.3倍以上,啤酒的6.0倍以上,葡萄酒的3.7倍以上[8,21-23]。氨基酸态氮是黄酒的重要质量指标,根据国家标准GB/T 13662—2008《黄酒》[1]规定,黄酒中氨基酸态氮的最低含量为0.3 g/L。表1所示为不同品牌黄酒中氨基酸态氮及氨基酸的含量。从表1中可知黄酒中氨基酸含量高的酒样其氨基酸态氮的含量相对也高,说明氨基酸态氮的含量能反应黄酒中氨基酸总体水平;氨基酸含氮量占氨基酸态氮比例越高,说明蛋白质的降解程度越大[24]。

表1 不同类型黄酒中氨基酸态氮及氨基酸的含量[24]Table1 Contents of amino acids and amino acids nitrogen in different types of Chinese rice wine
氨基酸对黄酒的色、香、味、形等各方面均有重要的贡献。首先,在陈酿时,氨基酸与低分子糖类物质通过氨基-羟基反应产生类黑精,是黄酒颜色的重要来源。氨基酸本身也是黄酒的重要风味物质,不同的氨基酸分别呈鲜、甜、苦、涩等味感,在一定程度上决定着黄酒的口味。同时,氨基酸也是黄酒中酵母代谢产生高级醇等挥发性风味物质的前体[24]。
同时,氨基酸和肽也赋予了黄酒一定的保健功能。黄酒中含有具有生理活性的肽,其中一些多肽具有抗氧化能力,能预防因自由基产生过多而引起的疾病,还含有降血压功能的降压肽[25-26]。谢广发等[3]成功鉴定出古越龙山加饭酒中的5种降血压活性肽(其氨基酸序列分别为Gln-Ser-Gly-Pro、Val-Glu-Asp-Gly-Gly-Val、Pro-Ser-Thr、Asn-Thr、Leu-Tyr)和另外一种降胆固醇活性肽(氨基酸序列为Cys-Gly-Gly-Ser)。黄酒中还富含天然非蛋白质氨基酸-γ-氨基丁酸(GABA),它是一种重要的抑制性神经递质,具有降血压、改善脑功能、延缓脑衰老等功效[8]。
2.2 蛋白质对黄酒浑浊的影响
黄酒酿造发展至今,消费者对其品质的要求越来越高,澄清的黄酒更容易被接受,然而非生物浑浊问题是黄酒行业仍然存在的亟待解决的问题之一。黄酒中蛋白质、多糖、多酚、金属离子等物质的存在使浑浊机理相对复杂,但根据目前的研究结果,认为蛋白质是引起黄酒非生物浑浊的主要原因[4-6]。
非生物浑浊的形成较为复杂,从形成机理来说,可分为蛋白质浑浊、铁浑浊和氧化浑浊[27]。此外,还有草酸钙浑浊[28]。其中蛋白质浑浊被认为是最主要、同时也是研究最多的浑浊成因。
2.2.1黄酒浑浊蛋白
杨国军等[7]通过对比不同酒样的稳定性发现,黄酒稳定性与酒中的某种特定蛋白有关,去除该类蛋白可提高黄酒稳定性,但作者并未指出这种特定蛋白是哪一类蛋白。表2所示为不同储存年份的绍兴花雕酒中蛋白质的分布、含量及稳定性。随着储存时间延长,总蛋白质含量降低,稳定性增加,说明蛋白质影响黄酒的非生物稳定性[7]。谭新勇[11]采用双向电泳技术及基质辅助激光解析电离飞行时间串联质谱法,鉴定出绍兴黄酒浑浊蛋白质主要有二聚-α-淀粉酶抑制剂、类燕麦蛋白、病程相关蛋白、几丁质酶Ⅱ和β-淀粉酶。其中来源于小麦的蛋白有二聚-α-淀粉酶抑制剂、类燕麦蛋白、病程相关蛋白、和几丁质酶Ⅱ,来源于水稻的蛋白有类燕麦蛋白和β-淀粉酶,由此推断,黄酒浑浊蛋白主要来源于大米和麦曲原料[11-12]。图1所示为浑浊蛋白双向电泳图。通过分析多种绍兴黄酒的浑浊蛋白发现,浑浊蛋白中含量最高的为二聚-α-淀粉酶抑制剂。

表2 不同储存年份绍兴花雕酒中蛋白质的分布、含量及稳定性[7]Table 2 Protein distribution, content and stability in Shaoxing Huadiao with different age

图1 浑浊蛋白双向电泳图谱[12] Fig.1 2-DE profiles of haze proteins in Chinese ricewine samples粗箭头()所指为二聚-α-淀粉酶抑制剂的蛋白点,细箭头(←)所指为类燕麦蛋白
二聚-α-淀粉酶抑制剂是α-淀粉酶抑制剂的二聚体,分子质量为24 kDa,在制曲过程中未被降解而保留至成品黄酒中[12]。王文蒙[29]研究发现,α-淀粉酶抑制剂具有较好的热稳定性和酸碱耐受性,当温度低于80 ℃时,α-淀粉酶抑制剂的活性几乎不受影响,在80 ℃下作用30 min,抑制活性仅损失10%左右,且在pH3~10之间抑制活性相对稳定。俞关松等[30]检测黄酒pH值约为3.50~4.50,由此推断在此pH值下α-淀粉酶抑制剂的活性较为稳定。因此,在黄酒煎酒的过程中α-淀粉酶抑制剂依然稳定存在,在瓶装黄酒运输、销售过程中受环境因素的影响慢慢析出,形成浑浊。小麦类燕麦蛋白具有高含量的半胱氨酸残基,半胱氨酸残基是形成二硫键的基础,而二硫键有助于蛋白之间的聚合,对浑浊的形成有重要影响[31]。
孙军勇等[12]对瓶装黄酒浑浊蛋白与酒体蛋白的氨基酸的分析表明,浑浊蛋白中谷氨酸、脯氨酸、天冬氨酸的含量较高,而在酒体蛋白中含量较高的前3位分别是谷氨酸、天冬氨酸和丝氨酸。其中谷氨酸、天冬氨酸、脯氨酸和丝氨酸的疏水值为分别为0、0、2.6和-0.3,谷氨酸、天冬氨酸和丝氨酸属于亲水性氨基酸,脯氨酸属于疏水性氨基酸[12]。可见,浑浊蛋白的疏水性高于酒体蛋白,导致其更容易从黄酒中析出。
2.2.2蛋白浑浊机理的研究
在成品黄酒中,蛋白质与酒中其它物质以胶体的状态存在,此时酒液澄清。在黄酒储存、运输过程中,由于外界环境的改变,以及酒体各组分物质的变化,胶体状态被破坏,出现浑浊现象。其中由蛋白质引起的浑浊可分为3种类型[27,32]:
(1)冷浑浊:冷浑浊属于可逆浑浊,是由β-球蛋白和醇溶蛋白与单体多酚结合形成可溶性多酚-蛋白质复合物而产生的。温度较高时,蛋白质和水结合形成氢键,酒液澄清;温度降低时,蛋白质与水形成的氢键断裂,然后与多酚结合形成浑浊。
(2)氧化浑浊:氧化浑浊属于不可逆浑浊,酒液长期放置时,在铁离子催化下,含巯基蛋白质被氧化,聚合成大分子蛋白,与聚多酚结合形成沉淀。
(3)热浑浊:热浑浊属于不可逆浑浊,是由高分子蛋白受热变性进而聚合析出形成的。热力作用破坏了高分子蛋白质的水化膜,蛋白质之间的电荷平衡被打破,进而形成沉淀。
有研究表明[27],黄酒浑浊的产生最初是由某些小分子物质相互作用,使酒液的平均粒度增大但<100 nm,出现“浑浊”现象,这些小分子物质在各种因素的作用下慢慢聚集,当酒液粒度>100 nm时,“浑浊”颗粒开始增大而沉降下来,形成沉淀。
研究者[33]在对葡萄酒的研究中模拟了浑浊蛋白-类甜蛋白(thaumatin-like proteins,TLPs)的三维结构,研究发现,引起浑浊的原因主要是高温环境下TLPs蛋白的三维结构被破坏,位于蛋白质内部的氨基酸侧链暴露,该侧链与其他蛋白质或葡萄酒中的其他组分结合,形成浑浊。蛋白质的结构决定其功能,不同的空间结构使蛋白质具有不同的理化性质和生理功能,在对TLPs蛋白的研究中发现,一些TLPs蛋白参与浑浊的形成,而另一些则不参与,这是由于TLPs蛋白不同异构体在结构上存在差异[33]。图2所示为TLPs蛋白不同异构体(F2/4JRU、I/4L5H)的三维结构。图2-A为两种异构体蛋白二级结构的重叠,可以看出F2/4JRU与I/4L5H的二级结构相似,只在两个环形区域有差异,其中图2-B为结构域1中的差异,图2-C为结构域2中的差异。此差异导致两个蛋白异构体具有不同的热稳定性,F2/4JRU为热不稳定蛋白,参与葡萄酒浑浊的形成,I/4L5H则较为稳定,不参与形成浑浊[33]。
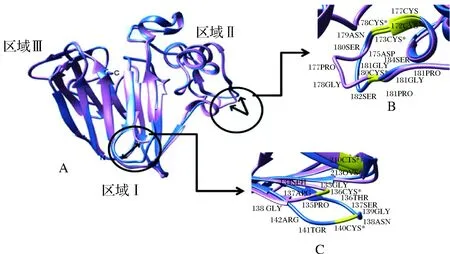
A: F2/4JRU与I/4L5H二级结构重叠,箭头所指为两个结构的差异所在;B: 结构域1中结构差异;C: 结构域2中结构差异图2 TLPs蛋白不同异构体(F2/4JRU、I/4L5H)的3D结构[33]Fig.2 The three-dimensional structures of TLP isoforms
黄酒浑浊的形成是否同样是浑浊蛋白-二聚-α-淀粉酶抑制剂的三维结构发生了变化,也可通过获取该蛋白的氨基酸序列模拟其三维结构来验证。
一些研究者[5,23,34]认为,黄酒中蛋白质与多酚结合形成缔合物是形成浑浊最主要的因素。林峰等[5]通过对酒脚成分的测定,以及对比酒脚与酒体成分的差异,推测高分子蛋白与单宁是形成浑浊的主要原因。江超等[10]通过分析不同年份的绍兴加饭酒发现,随着陈酿年份的增加,酒体中蛋白质和多酚的含量分别减少了55.23%和54.56%,由此推测蛋白与多酚结合是形成浑浊的主要原因。也有研究者[8]认为,黄烷醇类是影响黄酒浑浊的多酚物质。但目前为止,黄酒中多酚的种类尚不清楚,黄酒与多酚结合形成沉淀的机理也未得到证实。但黄烷醇参与葡萄酒蛋白浑浊的形成已经证实,图3所示为葡萄酒蛋白与黄烷醇结合形成浑浊的机制[35]。首先需要检测黄酒中多酚的种类,然后以葡萄酒蛋白与黄烷醇结合的机理为思路来验证黄酒蛋白与多酚结合是否是形成浑浊的原因。

图3 葡萄酒蛋白与黄烷醇结合形成沉淀的机理[35]Fig.3 Proteins precipitation via interaction with flavanols
2.2.3其他组分对浑浊形成的影响
瓶装黄酒浑浊物中,除蛋白质外,还包括多酚、多糖、金属离子铁等成分。这些成分对浑浊的形成也有影响,然而由于缺乏深入的研究,它们的影响机制尚不清楚。本文结合葡萄酒中非蛋白组分的研究进行论述,提供今后的研究方向,若能明确非蛋白组分在黄酒浑浊形成中的作用,可为解决非生物浑浊问题提供理论指导。
黄酒多酚主要来源于小麦,目前的一些文献普遍借鉴啤酒中的形成机制,认为主要是通过与蛋白质结合形成氢键,继而形成浑浊,但此观点在黄酒中是否正确还需实验的验证。
黄酒中的铁离子主要来源于原料及生产设备的腐蚀,其在氧化浑浊中起催化作用,目前普遍认为铁离子能促进黄酒浑浊的形成。黄酒中铁离子以Fe3+的形式存在,Fe3+与磷酸根离子结合形成带负电荷的磷酸铁胶体,此物质可与带正电荷的蛋白质等物质发生凝聚而形成浑浊[6,8]。
有关多糖对于葡萄酒浑浊的影响的研究表明[36-37],一些多糖能促进浑浊的形成,而一些低分子量的糖蛋白,例如甘露糖蛋白、阿拉伯半乳糖蛋白则抑制浑浊的形成。成品黄酒中含有大量的多糖,而且黄酒的陈酿时间较长,黄酒蛋白可能会与多糖结合形成糖蛋白,对浑浊的产生有一定的影响,且高度糖基化增加了蛋白分离纯化的难度。但黄酒中多糖的种类以及多糖与蛋白质之间的结合位点尚不清楚,需要进一步的研究。
2.2.4提高黄酒蛋白稳定性的方法
经发酵、陈酿后的原酒含有丰富的蛋白质,会引起瓶装黄酒的浑浊,因此,工业上通常在黄酒勾兑后、包装前,采用各种澄清技术去除导致黄酒浑浊的蛋白,常用方法有冷冻处理法、澄清剂法和微滤法等。
(1)冷冻处理法[8]:该方法将黄酒冷却至-6~-2 ℃,并维持3~5 d。冷冻处理法以黄酒的冷浑浊为依据,短时间内降低黄酒的温度,使酒液中的不稳定物质沉淀析出,该方法效果比较明显,但其耗电量较大,成本较高。
(2)澄清剂法[8]:澄清剂法是目前去除黄酒浑浊蛋白普遍使用的方法,常用的澄清剂有单宁、硅胶、皂土、PVPP、海藻酸钠和海藻酸丙二醇酯等。通常单一的澄清剂处理效果不佳,实际生产中一般采用混合澄清剂处理。许多研究表明[38-41],经过各种澄清方法处理之后,黄酒中蛋白质的含量明显降低,非生物稳定提高。但澄清剂法易受用量、酒体成分、pH等因素的影响,而且特异性不强,在去除黄酒浑浊蛋白的同时,也去除了酒体中与浑浊无关的其他蛋白,所以开发能够特异性去除二聚-α-淀粉酶抑制剂和类燕麦蛋白的澄清剂品种或者配方是今后的研究方向。
(3)微滤法[6]:微滤法是通过微滤膜两侧的压力差来筛分溶液中的颗粒和大分子物质,可直接过滤掉黄酒中易形成浑浊的大分子物质。谢广发[8]首次将0.8 μm的错流膜过滤与冷冻处理法相结合,显著提高了黄酒的非生物稳定性。
此外,向酒液中添加酶制剂降解浑浊蛋白的方法在啤酒和葡萄酒中均有应用,此方法可有效提高酒液的蛋白稳定性,且蛋白质降解的产物可以作为氮源被酵母吸收利用,例如在葡萄酒中添加AGP(一种曲霉菌肽酶)[42]。另外,还可以通过模拟浑浊蛋白的三维结构,研究其结构特异性,开发能够特异性水解这些蛋白质的蛋白酶,以防浑浊的形成[26]。如世界著名的酶制剂公司-荷兰皇家帝斯曼集团通过研究啤酒浑浊蛋白的结构和组成,从黑曲霉中分离得到了一种能够特异性降解浑浊蛋白的脯氨酸内切酶,而对啤酒中的其他蛋白特别是泡沫蛋白没有影响[43]。
3 展望
黄酒因其特有的营养保健功效越来越受到重视。随着蛋白质组学等技术手段的应用,以及对黄酒蛋白质研究的不断深入,将会有更多的功能性肽类物质被发现。同时,随着浑浊机理研究的进一步深入,黄酒行业一大难题-蛋白质浑浊,将会得到突破性的进展,最终被彻底解决。
今后,可以从以下几方面继续对黄酒蛋白进行深入研究:
(1)原料糯米中蛋白质提取与种类鉴定,并分析蛋白质的降解情况;
(2)从目前的报道中已知浑浊蛋白质的组成和来源,但对于瓶装黄酒酒体蛋白的组成尚不清楚。通过蛋白质组学技术分析瓶装黄酒酒体蛋白的组成,并与浑浊蛋白进行比较,有助于深入分析浑浊蛋白产生的原因;
(3)对各种澄清技术处理前后酒样中蛋白质种类和数量进行研究,分析各种澄清技术去除的蛋白种类和数量,为工艺生产中的应用提供理论指导;
(4)通过对浑浊蛋白研究分析可知,浑浊蛋白中来源于小麦的二聚α-淀粉酶抑制剂的含量最多,而黄酒酿造过程中小麦中的其他蛋白绝大部分被降解,通过研究二聚α-淀粉酶抑制剂的组成及结构特点,分析其未被蛋白酶降解以及未被其他工艺措施去除的原因,或许可以找到特异性去除黄酒浑浊蛋白的方法;
(5)研究黄酒浑浊中的非蛋白组分,如多糖的种类和数量等,分析这些组分对蛋白质浑浊形成的影响。
[1]GB/T 13662-2008, 黄酒 [S]. 北京: 中国标准出版社, 2008.
[2]倪赞. 中国黄酒保健功能的研究 [D]. 杭州: 浙江大学, 2006.
[3]谢广发. 黄酒的功能性成分与保健功能 [J]. 酿酒, 2008, 35(5):76-77.
[4]谢广发, 孟中法, 周建弟. 黄酒蛋白质的测定及其沉淀原因探讨 [J]. 酿酒科技, 2002, (03):64-68.
[5]林峰, 白少勇, 邹慧君, 等. 黄酒蛋白质沉淀 [J]. 酿酒科技, 2005(9):69-72.
[6]詹汉林, 肖招燕, 韩珍, 等. 黄酒中蛋白质沉淀的研究进展 [J]. 食品研究与开发, 2014, 35(12):130-134.
[7]杨国军, 俞关松, 尉冬青. 黄酒中蛋白质分布及含量与酒质稳定性关系的研究 [J]. 中国酿造, 2005,24(10):47-49.
[8]谢广发. 黄酒酿造技术(第2版) [M]. 北京: 中国轻工业出版社, 2016:15;185-189.
[9]胡普信. 中国黄酒的现状思考 [J]. 中国酒, 2013(4):50-54.
[10]江超, 林峰, 邹慧君, 等. 陈年绍兴黄酒的成分分析与品质鉴定 [J]. 食品与发酵工业, 2009, 35(10):119-123.
[11]谭新勇. 黄酒非生物混浊的初步研究 [D]. 无锡: 江南大学, 2013.
[12]孙军勇, 樊世英, 谢广发, 等. 绍兴黄酒混浊蛋白的分离鉴定及其氨基酸组成、二级结构分析 [J]. 食品与发酵工业, 2016, 42(2):1-6.
[13]张波, 余培斌, 陆健. 绍兴黄酒熟麦曲制曲过程的宏蛋白质组学研究 [J]. 中国酿造, 2014, 33(2):101-104.
[14]ODA K, KAKIZONO D, YAMADA O, et al. Proteomic analysis of extracellular proteins fromAspergillusoryzaegrown under submerged and solid-state culture conditions[J]. Applied and Environmented Microbiology, 2006,72(5):3 448-3 457.
[15]孔令琼, 管政兵, 陆健, 等. 绍兴黄酒成品麦曲中微生物胞外酶的双向电泳技术的建立 [J]. 食品与生物技术学报, 2011, 30(3):453.
[16]孔令琼. 黄酒麦曲浸提液的宏蛋白质组学研究 [D]. 无锡: 江南大学, 2011.
[17]程秀秀. 黄酒组成成分与原料糯米营养成分的相关性研究 [D]. 杭州: 浙江农林大学, 2014.
[18]陈金斌, 芮鸿飞, 方佳宁, 等. 糯米蛋白质与黄酒氨基酸的相关性分析 [J]. 酿酒科技, 2017(4): 51-56.
[19]代钰, 丁文平, 王毅梅, 等. 稀碱法分离工艺对糯米中蛋白质提取率的影响 [J]. 中国粮油学报, 2011, 26(04):1-5.
[20]ROWE J D. Yeast protein release during fermentation and aging in a model wine [D]. Oregon: Oregon State University, 2008.
[21]范怀德, 乔自林. 黄酒营养价值的研究 [J]. 西北民族学院学报(自然科学版), 2000, 21(2):47-49;61.
[22]周建弟. 浅谈黄酒中的氨基酸及其含量的控制 [J]. 酿酒科技, 2002(4):73-74.
[23]白少勇. 黄酒中蛋白质的沉淀机理与处理方法的研究 [D]. 杭州: 浙江大学, 2005.
[24]潘慧青. 黄酒中氨基酸态氮的来源及酿造工艺的影响 [D]. 无锡: 江南大学, 2015.
[25]孟如杰. 黄酒中抗氧化活性物质的研究 [D]. 无锡: 江南大学, 2008.
[26]倪莉, 吕旭聪, 黄志清, 等. 黄酒的生理功效及其生理活性物质研究进展 [J]. 中国食品学报, 2012, 12(3):1-7.
[27]林峰, 白少勇. 黄酒非生物浑浊和沉淀的特点与解决方法 [J]. 酿酒科技, 2005(10):68-74.
[28]谢广发, 沈斌, 胡志明, 等. 黄酒中的另一种沉淀—草酸钙沉淀 [J]. 酿酒, 2011, 38(3):26-28.
[29]王文蒙. α-淀粉酶抑制剂的提取、分离及性质研究 [D]. 天津: 天津商业大学, 2010.
[30]俞关松. 黄酒pH值指标范围的探讨及超标的预防控制 [J]. 食品与发酵工业, 2002, 28(1):76-78.
[31]刘威. 小麦Avenin-like b蛋白对小麦加工品质参数的影响 [D]. 武汉: 华中科技大学, 2014.
[32]权美平, 田呈瑞. 黄酒混浊沉淀的原因及防止措施 [J]. 晋东南师范专科学校学报, 2004, 21(2):47-49.
[33]MARANGON M, VAN SLUYTER S C, WATERS E J, et al. Structure of haze forming proteins in white wines: vitis vinifera traumatic-like proteins [J]. Plos One, 2014, (9):1-21.
[34]蔡小云. 黄酒乙醇—浊度法及其机理研究 [D]. 杭州: 浙江大学, 2008.
[35]BENUCCI I. Removal of unstable proteins from white wine by immobilized acid protease [D]. Viterbo: Tuscia, 2016.
[36]FERREIRA R B, PICARRA-PEREIRA M A, MONTEIRO S, et al. The wine proteins [J]. Food Science and Technology, 2002(12):230-239.
[37]DUFRECHOU M, DOCO T, PONCET-LEGRANT C, et al. Protein/polysaccharide interactions and their impact on haze formation in white wines [J]. Agricultural and Food Chemistry, 2016, 64:6 752-6 761.
[38]杨国军. 澄清剂在预防黄酒非生物混浊中的应用 [J]. 山东食品发酵, 2005(1):23-25.
[39]李艳敏, 赵树欣. 不同酒类澄清剂的澄清机理与应用 [J]. 中国酿造, 2008,27(1):1-5.
[40]陈珊, 曾庆梅. 不同澄清剂协同超高压处理对黄酒的稳定性研究 [J]. 安徽农业科学, 2015, 43(11):269-273.
[41]樊世英, 孙军勇, 谢广发, 等. 澄清剂对黄酒混浊蛋白去除效果的研究 [J]. 食品工业科技, 2015, 36(8):167-170;177.
[42]VAN SLUYTER S C, MCRAE J M, FALCONER R J, et al. Wine protein haze: mechanisms of formation and advances in prevention [J]. Agricultural and Food Chemistry, 2015, 63:4 020-4 030.
[43]FINN J. Investigation of the impact of proline specific endoproteinase on malt haze-active and foam-active proteins[C]. Proceedings of the Australian Barley Technical Symposium. Fremantle, Western Australia, 26-29 August 2007.
