支持柱直径对紫萼龙吐珠形态特征和生物量分配的影响
2018-04-10朱锦心翁殊斐冯嘉仪范体凤
朱锦心,翁殊斐,李 灿,冯嘉仪,范体凤
(华南农业大学 林学与风景园林学院,广东 广州 510642)
木质藤本植物是热带森林的重要结构和功能组分[1-2],因其具有占地面积小、生长速度快、抗逆性强等特点,成为提高城市绿化覆盖率、丰富绿化层次的不可或缺的垂直绿化材料[3]。木质藤本植物在生长过程中缺乏直立生长能力,需要攀援其他植物或物体获得支持,以拓展空间获取更多的资源[4],由于受支持物资源的限制,加之生境的时空变化十分剧烈,木质藤本植物在形态和生物量分配等方面常发生明显变化,表现出较大的可塑性[5-6]。
支持物的不同形式决定着藤本植物寻觅外界资源的能力,因此支持物的有无及其存在状态对藤本植物的生长和适应性有着重要影响。木质藤本植物围绕直立的外界支持物的成功缠绕显示出其行为的稳定性与其茎的螺旋结构息息相关,研究发现,攀援茎的螺旋结构如螺旋波长(wavelength)、曲率(curvature)和扭转率(torsion)会随着支持物直径大小而发生变化[7-8]。目前对草质藤本植物与支持物[9]、森林群落中藤本植物与支持木关系[10-11]的研究已经较为深入,还未见有关城市园林中木质藤本与支持物关系的报道。因此,本研究以南亚热带地区常见的缠绕类木质藤本植物紫萼龙吐珠(Clerodendrumspeciosum)为材料,通过对其在不同直径支持柱上主茎和分枝水平形态特征、生物量分配的差异研究,选用扭转率作为衡量紫萼龙吐珠缠绕效率的指标,揭示其在不同直径支持柱条件下的适应能力及策略,以期为城市园林垂直绿化中特定木质藤本景观形式(如篱垣式和栅栏式)的快速攀援及苗木栽培提供理论依据和技术指导。
1 材料与方法
1.1 研究地概况
研究地点位于广东省广州市华南农业大学苗圃内,该地位于北纬23°09′28″,东经113°21′07″。地理上属南亚热带季风气候区,全年光照充足,雨量充沛,年平均气温为21.4~21.9 ℃,11月下旬至2月中旬有时出现霜冻;年平均降水量为1 623.6~1 899.8 mm,年平均日照时数为1 820~1 960 h。
1.2 试验材料
选择紫萼龙吐珠1年生扦插苗为研究对象。紫萼龙吐珠为马鞭草科Verbenaceae赪桐属Clerodendrum多年生常绿缠绕类木质藤本植物,是龙吐珠C.thomsonae和美丽赪桐C.splendens的杂交种,以观花为主,聚伞花序腋生或顶生,花色艳丽,小花红色,5瓣,花冠易脱落,但萼片长久留存,成为主要观赏部位,观赏期为全年,盛花期9~12月,恰逢广州地区少花季节[12]。
1.3 试验设计
从长势良好、无病虫害的1年生紫萼龙吐珠扦插苗中,选择120株大小和生长状况一致的幼苗,移栽在深20 cm、直径25cm的花盆中,移栽土壤为壤土,参考前人研究[13]选用直径分别为8,16,24 mm的上下端粗细一致的竹竿,同地平面的角度均为90°,并以无支持柱作为对照(CK)。每种直径支持柱处理10株幼苗,设置3次重复。在试验初期人为将植株引向支持柱,试验期间统一水肥管理。试验于2016年5月10日开始,在5月17日、5月24日、5月31日、6月7日、6月14日进行5次生长动态记录,测量指标包括主茎长、节间长和总叶片数;最后按照随机采样法从各重复中选择5株,于6月14日进行形态参数和生物量参数等指标的测定。
1.4 测定指标与方法
1.4.1形态参数按主茎和分枝水平记录不同直径支持柱下植株的叶片数、一级分枝数和二级分枝数。用卷尺或直尺(精确度为1 mm)测量植株的主茎(分枝茎)长、叶柄长度、节间长度,用游标卡尺(精确度为0.1 mm)测定主茎和分枝水平的地径,用Win Folia 2008软件测定叶面积。分枝数按照分枝茎的自然长度不小于10 cm,且有2片以上叶片计算,叶片数以叶片完全展开为准,将植株按其分枝发育顺序的离心法分级,即根茎长出的作为一级分枝,一级分枝上的侧枝为二级分枝(二级分枝相遇处为一级分枝)[14],分枝率=分枝数/相对应的茎长度。比茎长(Specific stem length,SSL)=茎长/茎干质量;比叶柄长(Specific petiole length,SPL)=叶柄长/叶柄干质量;比叶面积(Specific leaf area,SLA)=总叶面积/叶片总干质量。

1.4.3生物量参数将每个处理分为根、茎、叶、叶柄,于80 ℃下烘48 h至恒质量,用电子天平(精确度为0.000 1 g)按照主茎水平和分枝水平分别称量各组分干质量。计算各自的根冠比(Root-shoot ratio,RSR)=根干质量/地上部分总干质量;根生物量比(Root biomass rate,RBR)=根干质量/植株总干质量;茎生物量比(Stem biomass rate,SBR)=茎干质量/植株总干质量;叶生物量比(Leaf biomass rate,LBR)=叶干质量/植株总干质量。
1.4.4表型可塑性指数根据Valladares等[15]的方法计算每个指标的表型可塑性指数[(不同处理下某一变量的最大值-其最小值)/其最大值]。平均形态可塑性和生物量可塑性指数是3个直径支持柱处理植株各形态和生物量指标可塑性指数的平均值,反映紫萼龙吐珠对不同直径支持柱的相对平均适应能力。
1.5 数据分析
采用Excel 2007软件进行数据处理,运用SPSS 19.0统计软件进行单因素方差分析,采用Duncan’s检验法进行差异显著性分析,并采用Origin 8.5软件制图。所有的数据在进行分析之前都进行正态性和方差齐性检验,当不满足时对数据进行对数转换。
2 结果与分析
2.1 支持柱直径对紫萼龙吐珠生长动态的影响
不同直径支持柱下紫萼龙吐珠的生长动态见图1。

图1 不同直径支持柱下紫萼龙吐珠的生长动态Fig.1 Growth dynamics of Clerodendrum speciosum under external supports with different diameters
图1显示,在紫萼龙吐珠的攀援过程中,不同直径支持柱对植株的生长动态具有显著影响(P<0.05)。较小直径支持柱下植株的主茎长大于较大直径支持柱,直径8 mm支持柱上紫萼龙吐珠节间长的生长速度在试验开始后7 d和试验结束前7 d均表现出加快,表明在较小直径支持柱下紫萼龙吐珠主要通过增大主茎长和节间长的方式进行有效的自主攀援。直径24 mm支持柱上植株的叶片数于5月31日开始多于直径16 mm支持柱,而其主茎长依然小于直径16 mm支持柱,显示紫萼龙吐珠在较大直径支持柱上通过增加分枝数的方式提高对外界合适支持物的寻觅能力。无支持柱组植株的各指标生长速度明显小于其他3组处理,结果显示支持物的存在可以帮助紫萼龙吐珠生长得更快,对其寻觅支持物和光照资源有一定的帮助。
2.2 支持柱直径对紫萼龙吐珠形态参数的影响
由表1可见,紫萼龙吐珠在直径8,16,24 mm支持柱下的螺旋缠绕等级分别为2、3、1,显示支持柱直径大小可以影响植株的自主螺旋缠绕效率,当支持柱直径为16 mm时植株的自主缠绕效果最有效,而其产生的一级分枝数和一级分枝率均显著小于其他处理(P<0.05)。
由表1可见,支持柱直径的大小通过影响植株的攀援状况而对植株的生长施加影响,在主茎水平的形态特征方面,从主茎长和叶片数指标看,直径16 mm支持柱上的植株均显著大于直径24 mm支持柱上的植株,与直径8 mm支持柱上的植株无显著差异(P>0.05),无支持物组植株显著小于其他3种直径支持柱上的植株;而从叶柄长指标看,直径16 mm支持柱上的植株显著小于直径24 mm支持柱上植株。结果显示,获得有效攀援的植株能更好地伸展主茎,而未能有效攀援的则通过增大主茎叶柄长、增加分枝数的方式寻觅新的、合适的支持物。

表1 不同直径支持柱下紫萼龙吐珠主茎水平的形态参数Table 1 Stem morphological parameters of Clerodendrum speciosum under external supports with different diameters
注:同列数据后标不同小写字母表示差异显著(P<0.05)。
Note:Different lowercase letters in each column mean significant difference (P<0.05).
在分枝水平的形态特征方面(图2),直径16 mm支持柱上的植株分枝叶片数、节间长、分枝茎长均与对照组间无显著差异(P>0.05),显示在遇到合适粗细的支持柱时,缠绕型木质藤本植物的生长将集中在主茎上,从而弱化了其分枝生长。缠绕情况较好的植株(直径16 mm支持柱)的分枝叶片数显著少于其他2个直径支持柱处理(P<0.05),缠绕情况较差的植株(直径24 mm支持柱)的形态参数均显著大于直径16 mm支持柱上的植株。显示植株的自主攀援有效性越差,其分枝水平的形态参数会越大。
2.3 支持柱直径对紫萼龙吐珠生物量及其分配的影响
表2显示,在主茎水平上,除了总生物量之外,不同直径支持柱下植株的各组分生物量和生物量比均没有显著差异(P>0.05),无支持柱组植株的根冠比显著大于有支持柱的植株(P<0.05),表明支持柱的存在更多地增加了植株地上部总生物量。在主茎和分枝水平上,直径8 mm支持柱上植株的总生物量均显著高于直径16 mm支持柱上的植株。在分枝水平上,攀援较好的植株(直径为16 mm支持柱的植株)的茎生物量、总生物量显著小于其他2个直径处理的植株,且随缠绕等级的下降,茎生物量、叶柄生物量和茎生物量比都增加。结果显示,随着植株扭转率下降,攀援外界支持柱的有效性降低,植株会将生物量更多地分配到分枝茎和叶柄上,达到进一步搜寻到支持物的目的。

图柱上标不同小写字母表示处理间差异显著(P<0.05)Different lowercase letters show significant difference among treatments at P<0.05 level

生物量参数Biomassparameter支持柱直径/mmExternalsupportdiameter8 16 24 CK 总生物量/(g·株-1)Totalbiomass8.99±1.78a6.37±1.01bc7.50±1.38ab3.78±0.87c根冠比Root⁃shootratio0.13±0.07b0.10±0.01b0.12±0.03b0.43±0.12a主茎水平Stemlevel根生物量比Rootbiomassrate0.11±0.05b0.09±0.01b0.11±0.02b0.30±0.06a茎生物量比Stembiomassrate0.24±0.04a0.22±0.06a0.24±0.04a0.18±0.04b叶生物量比Leafbiomassrate0.62±0.03a0.65±0.06a0.62±0.03a0.50±0.02b茎生物量/(g·株-1)Stembiomass0.50±0.19a0.07±0.04b0.66±0.28a0.03±0.02b叶生物量/(g·株-1)Leafbiomass1.69±0.65a0.31±0.15b1.27±0.02a0.75±0.30ab分枝水平Branchlevel叶柄生物量/(g·株-1)Petiolebiomass0.07±0.03b0.01±0.00b0.10±0.02a0.06±0.02b总生物量/(g·株-1)Totalbiomass2.26±1.03a0.38±0.16b2.04±1.01a0.84±0.33b茎生物量比Stembiomassrate0.25±0.09a0.19±0.09b0.43±0.21a0.04±0.01c叶生物量比Leafbiomassrate0.71±0.11c0.79±0.12b0.49±0.22c0.89±0.05a
注:同行数据后标不同小写字母表示差异显著(P<0.05)。
Note:Different lowercase letters in each row mean significant difference (P<0.05).
2.4 支持柱直径对紫萼龙吐珠表型可塑性指数的影响
表3和表4显示,在主茎水平上,紫萼龙吐珠的形态参数中,主茎长和一级分枝数表型可塑性指数均较高,在0.65以上;而其生物量参数中,仅根冠比和根生物量比的表型可塑性指数较高,分别为0.767,0.700,主茎水平的平均形态可塑性稍高于平均生物量可塑性。分枝水平上,形态参数中,紫萼龙吐珠分枝的叶片数、二级分枝数、比茎长和比叶柄长的表型可塑性指数均较高;而生物量参数中,除了叶生物量比的表型可塑性指数低于0.65外,其他参数的表型可塑性均较高。总体而言,在分枝水平上,紫萼龙吐珠的平均生物量可塑性高于平均形态可塑性。由此可见,不同直径支持柱对紫萼龙吐珠主茎水平的平均形态可塑性和分枝水平的平均生物量可塑性影响较大,而分枝水平相对较高的平均形态和生物量可塑性指数意味着其具有较强的寻觅外界支持物的能力,植株能够有效地利用有限资源,维持较高的生长速率。
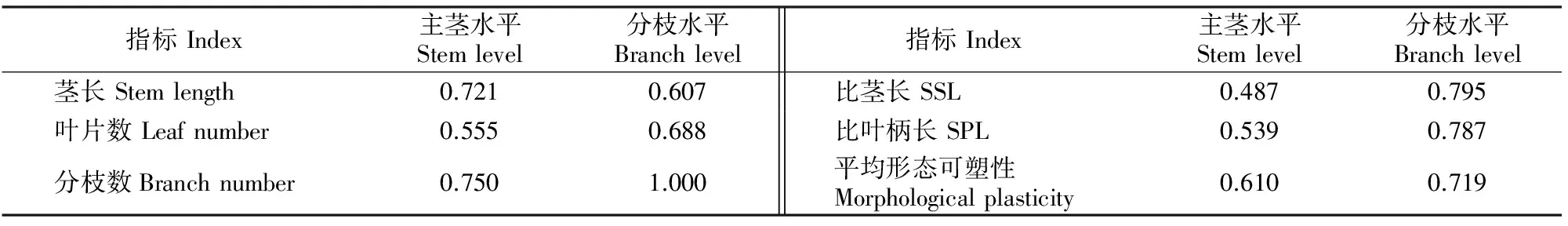
表3 不同直径支持柱下紫萼龙吐珠形态参数的表型可塑性指数Table 3 Index of morphological plasticity for Clerodendrum speciosum under external supports with different diameters
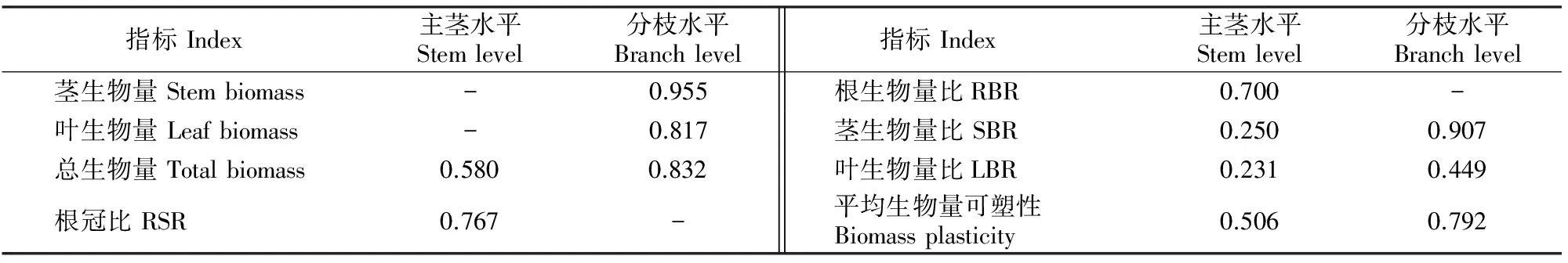
表4 不同直径支持柱下紫萼龙吐珠生物量参数的表型可塑性指数Table 4 Index of biomass plasticity for Clerodendrum speciosum under external supports with different diameters
3 讨论与结论
支持物作为藤本植物攀援生长所必需的一种资源,其有效性是制约藤本植物生长和觅食的重要因素,支持物的有无及其性状实质上是改变了藤本植物的光环境。缠绕类藤本植物攀援外界支持物呈现螺旋管状结构,其成熟的茎在直径均衡的支持物上往往拥有统一的扭转率[7]。本研究结果显示,各直径支持柱上紫萼龙吐珠的扭转率τ不同,且有显著差异,因此考虑用扭转率量化其自主攀援能力是可行的。在直径为16 mm支持柱上的紫萼龙吐珠具有较高的螺旋缠绕能力,且植株通过增加叶片数和主茎长提高自主攀援能力。无支持柱组植株的根冠比显著大于有支持柱的植株,表明支持柱的存在更多地是增加了植株地上部生物量。本研究结果显示,直径为8和16 mm的支持柱均较有利于紫萼龙吐珠的攀援生长,这与前人有关森林群落[16-17]的研究结果类似,其原因可能是由于缠绕类木质藤本植物作为一种比较原始的攀援类型,其通常要求中小径级支持木。由生长动态分析可见,支持柱的存在对紫萼龙吐珠的生长具有显著的促进作用,这一结论是否适合其他木质藤本植物还有待进一步研究。
适应是植物个体对变化环境所作出的形态和生理的调整,植物常通过表型改变来响应环境的变化,所以表型可塑性常被认为是植物适应环境的一种积极生长策略[18]。植物通过其个体或构件形态和生物量分配格局的改变,提高对外界资源最大限度的利用,各构件的生长速度可以解释植物在生境中采用的生活策略和资源利用效率[19]。陶建平等[13]研究发现,卷须类草质藤本植物苦瓜在不同直径支持物上节间的生长速度较快。本试验研究结果显示,直径8 mm支持柱上紫萼龙吐珠各指标均增长较快,而各处理节间长的生长速度相对较慢,这与前人研究结果不同,可能是由于缠绕类木质藤本植物的可塑性相比具有特化器官(卷须)的藤本植物小之故。本研究结果显示,紫萼龙吐珠主茎水平的外部形态和分枝水平的各器官生物量分配在异质环境中具有更大的可变性。
综上所述,在不同直径支持柱条件下,紫萼龙吐珠通过形态特征、生物量及其分配的改变,能促进其有效地自主攀援生长和寻觅外界支持物。对主茎和分枝水平的分析结果显示,紫萼龙吐珠主要通过增大主茎叶柄长、增加分枝数、比分枝茎长和比分枝叶柄长的方式提高对外界支持物的寻觅能力。对于生物量及其分配方面,不同直径支持柱对于植株分枝水平的影响较大;从主茎水平看,直径为8 mm支持柱有利于紫萼龙吐珠的生物量积累,并且植株的主茎长、节间长和叶片数生长更快,而直径16 mm支持柱更有利于植株的自主有效攀援生长,因此在实际园林应用中选取直径在8~16 mm的支持柱供其攀援更为合适。
[参考文献]
[1]Schnitzer S A,Bongers F,Burnham R J,et al.Ecology of lianas [M].UK:Wiley Blackwell,2015:1-481.
[2]Muthumperumal C,Parthasarathy N.Diversity,distribution and resource values of woody climbers in tropical forests of southern Eastern Ghats,India [J].Journal of Forestry Research,2013,24(2):365-374.
[3]袁喆,翁殊斐,王琳琅.广州园林绿地木质藤本景观美景度评价 [J].西北林学院学报,2015,30(4):301-306.
Yuan Z,Weng S F,Wang L L.Scenic beauty estimation of liana landscape in Guangzhou green open spaces [J].Journal of Northwest Forestry University,2015,30(4):301-306.
[4]樊星,王大伟,刘金平.支持物对攀援葎草分枝形态塑性和繁殖对策影响的性别差异 [J].生态学杂志,2015,34(6):1559-1564.
Fan X,Wang D W,Liu J P.Gender differences in morphological plasticity and reproductive strategy ofHumulusscandensin response to different external supports [J].Chinese Journal of Ecology,2015,34(6):1559-1564.
[5]吴涛,耿云芬,柴勇,等.三叶爬山虎叶片解剖结构和光合生理特性对3种生境的响应 [J].生态环境学报,2014,23(10):1586-1592.
Wu T,Geng Y F,Chai Y,et al.Response of leaf anatomical structure and photosynthesis characteristics ofParthenocissushimalayanato three habitat types [J].Ecology and Environmental Sciences,2014,23(10):1586-1592.
[6]钟章程.攀援植物行为生态学的理论与研究方法 [M].北京:科学出版社,2005:19-26.
Zhong Z C.Theory and research method on the behavioral ecology of climbing plant [M].Beijing:Science Press,2005:19-26.
[7]Isnard S,Silk W K.Moving with climbing plants from Charles Darwin’s time into the 21st century [J].American Journal of Botany,2009,96(7):1205-1221.
[8]Silk W K,Holbrook N M.The importance of frictional interactions in maintaining the stability of the twining habit [J].American Journal of Botany,2005,92(11):1820-1826.
[9]赵则海,余曼玲,陈雄伟,等.不同攀援角度裂叶牵牛叶片金属元素含量变化及其相关性分析 [J].生态环境学报,2009,18(2):669-673.
Zhao Z H,Yu M L,Chen X W,et al.Effects of different climbing angles on metallic element contents and their correlations in leaves ofPharbitisnil[J].Ecology and Environmental Sciences,2009,18(2):669-673.
[10]Garbin M L,Carrijo T T,Sansevero J B B,et al.Subordinate,not dominant,woody species promote the diversity of climbing plants [J].Perspectives in Plant Ecology,Evolution and Systematics,2012,14(4):257-265.
[11]Addo-Fordjour P,Rahmad Z B,Shahru A M S.Effects of human disturbance on liana community diversity and structure in a tropical rainforest,Malysia:implication for conservation [J].Journal of Plant Ecology,2012,5(4):391-399.
[12]朱锦心,冯嘉仪,翁殊斐,等.不同浓度IBA、NAA对紫萼龙吐珠扦插生根的影响 [J].亚热带植物科学,2016,45(3):285-289.
Zhu J X,Feng J Y,Weng S F,et al.Effect of different concentrations of IBA and NAA onClerodendrumspeciosumcutting [J].Subtropical Plant Science,2016,45(3):285-289.
[13]陶建平,钟章成,黄林.外界支持物直径对苦瓜生长和觅食行为的影响 [J].生态学杂志,2003,22(1):9-13.
Tao J P,Zhong Z C,Huang L.Effect of support diameter on growth and foraging behavior ofMomordicacharantia[J].Chinese Journal of Ecology,2003,22(1):9-13.
[14]李得禄,廖空太,严子柱,等.不同类型防护林中樟子松的构型特征分析 [J].中国沙漠,2008,28(1):113-118.
Li D L,Liao K T,Yan Z Z,et al.Analysis on branching architectures characteristics ofPinussylvestrisvar.mongolicain different shelterbelts [J].Journal of Desert Research,2008,28(1):113-118.
[15]Valladares F,Wright S J,Lasso E,et al.Plastic phenotypic response to light of 16 congeneric shrubs from a Panamanian rainforest [J].Ecology,2000,81(7):1925-1936.
[16]袁春明,刘文耀,杨国平.哀牢山中山湿性常绿阔叶林木质藤本植物的多样性与空间分布 [J].生物多样性,2015,23(3):332-340.
Yuan C M,Liu W Y,Yang G P.Diversity and spatial distribution of lianas in a mid-montane moist evergreen broad-leaved forest in the Ailao Mountains,SW China [J].Biodiversity Science,2015,23(3):332-340.
[17]胡亮,李鸣光,李贞.中国种子植物区系中的藤本多样性 [J].生物多样性,2010,18(2):212-221.
Hu L,Li M G,Li Z.The diversity of climbing plants in the spermatophyte flora of China [J].Biodiversity Science,2010,18(2):212-221.
[18]周浩然,徐明,于秀波,等.生物适应性和表型可塑性理论研究进展 [J].西北农林科技大学学报(自然科学版),2014,42(4):215-220,228.
Zhou H R,Xu M,Yu X B,et al.Study on acclimation and phenotypic plasticity issues [J].Journal of Northwest A&F University(Nat Sci Ed),2014,42(4):215-220,228.
[19]Franzese J,Ghermandi L.Early competition between the exotic herbRumexacetosellaand two native tussock grasses with different palatability and water stress tolerance [J].Journal of Arid Environments,2014,106(7):58-62.
